№1, 2007 г.
№1, 2007 г.
ЛАУРЕАТЫ НОБЕЛЕВСКОЙ ПРЕМИИ 2006 ГОДА По химии - Р. Корнберг
Нобелевская премия по химии за 2006 год присуждена Роджеру Д. Корнбергу “за фундаментальные исследования молекулярных основ транскрипции у эукариот”.
Транскрипция - это процесс матричного синтеза РНК на ДНК, который осуществляется ферментами РНК-полимеразами. У эукариот в него вовлечены несколько сложных олигомерных белковых комплексов: РНК-полимераза, общие факторы транскрипции и белковый комплекс медиатора. Перечисленные комплексы вместе со специфическими факторами транскрипции (активаторами) инициируют синтез РНК, начиная с промотора соответствующего гена в ДНК.
Роджер Корнберг (Roger D.Kornberg) родился 24 апреля 1947 г. в Сент-Луисе, штат Миссури, США. В 1967 г. окончил Гарвардский университет, в 1972-м получил докторскую степень в Станфордском университете (штат Калифорния). Сейчас, будучи профессором медицины, руководит отделом структурной биологии в медицинской школе того же университета.Корнберг - член Национальной академии наук и Американской академии искусств и наук; лауреат премии Уэлча - высшей награды по химии в США; лауреат премии Шарля Леопольда Майера, присуждаемой за биомедицинские исследования Французской академией наук.
Корнберг, как отмечено Нобелевским комитетом, внес наиболее существенный вклад в определение структуры компонентов транскрипционного комплекса, в установление динамики процессов, протекающих в его недрах, и регуляторных механизмов, которые ускоряют, замедляют или прерывают инициацию транскрипции. Многолетние (около 30 лет) исследования с использованием совершенной биохимической техники в сочетании с рентгеноструктурным анализом комплексов РНК-полимеразы II дрожжей с другими компонентами системы послужили основанием для получения картины, описывающей работу активного центра этого фермента, а также его взаимодействия с другими белками.
Исследование транскрипции у эукариот ведет начало с работы С.Б.Уэйсса и Л.Гладстона (1959). Однако очистка РНК-полимеразы эукариот оказалась очень сложным делом. Значительно быстрее развивались исследования транскрипции у бактерий. И уже в 1965 г. за успехи в изучении этого процесса была присуждена Нобелевская премия по физиологии и медицине Ф.Жакобу, Ж.Моно и А.Львову. Всего через несколько лет было установлено, что транскрипционная машина эукариот много сложнее: она включает три РНК-полимеразы - I, II и III. При этом все гены, кодирующие белки, транскрибируются РНК-полимеразой II. Однако in vitro она проявляла очень низкую активность в отношении генов, содержащих промоторы, которые служат сигналом для инициации синтеза РНК. С начала 70-х годов сформировались задачи, которые в то время казались неосуществимыми: выявить, очистить и охарактеризовать все компоненты транскрипционного комплекса; реконструировать модель транскрипции in vitro; установить все стадии матричного процесса на атомном уровне. В эту гигантскую работу включились несколько групп ученых, главным образом в США. Значительный вклад внесла группа Р.Редера, работавшая в Рокфеллеровском университете: им первым удалось осуществить инициацию транскрипции РНК-полимеразой II с вирусного промотора. Кроме того, были идентифицированы белковые факторы, получившие наименование общих, или универсальных факторов, так как они необходимы для транскрипции всех генов эукариот, кодирующих белки.
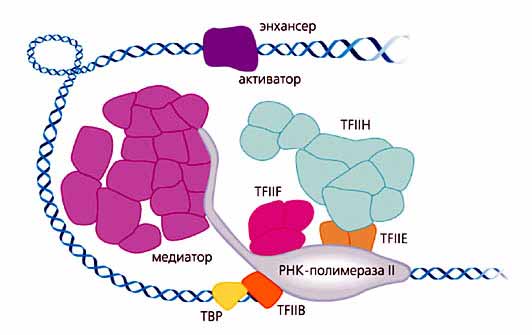
Свою первую работу Корнберг провел и завершил в начале 70-х годов в Кембридже совместно с Ф.Криком и А.Клугом. Оказалось, что основным элементом хроматина служит нуклеосома - гистоновый октамер, который образует комплекс с 200 парами оснований ДНК. После возвращения в середине 70-х в Станфорд (Калифорния) Корнберг начал исследовать регуляцию трансрипции у эукариот. В качестве модели он избрал пекарские дрожжи. В первых работах выяснилось, что очищенная РНК-полимераза II и пять общих факторов транскрипции (рис.1) - TFIIB, E, F, H и TBP - способны поддерживать лишь очень низкий, базовый уровень процесса. Добавление в систему специфических факторов транскрипции, взаимодействующих с промотором или энхансером и необходимых для узнавания определенного гена, не изменяло эту картину. Тщательное исследование клеточного экстракта привело к неожиданному открытию и очистке мультибелкового комплекса, названного медиатором (см. рис.1). Этот комплекс включал 20 различных белков. Роль медиатора, как выяснили Корнберг и сотрудники, состояла в переносе сигнала (как положительного, так и отрицательного) от специфического фактора транскрипции, связанного с энхансером, к РНК-полимеразе II. Медиатор оказался недостающим звеном, открытие которого объяснило непонятные ранее факты и сделало возможным реконструкцию транскрипционной машины in vitro.
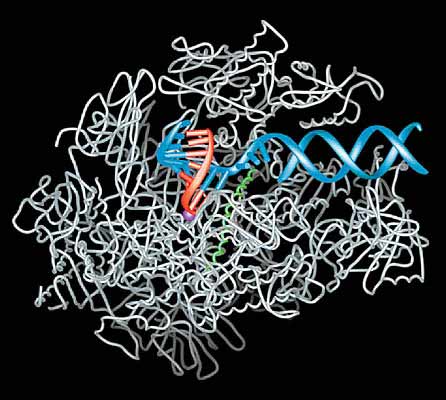
Рис. 1. Схематическое изображение комплекса инициации транскрипции ДНК. В составе комплекса приведены общие факторы транскрипции (TFIIB, E, F, H и TBP), РНК-полимераза II, медиатор и специфический фактор транскрипции, связанный с энхансером - последовательностью, довольно удаленной от промотора и регулирующей его.В целом транскрипционный комплекс состоит примерно из 60 взаимодействующих белков и имеет массу, превышающую 3МДа. Это создавало трудности для молекулярного исследования полного комплекса. Поэтому рентгеноструктурные исследования были направлены на полимеразу, своеобразную платформу (так назвал фермент сам Корнберг), вокруг которой собираются все элементы транскрипционной машины. В течение нескольких лет были проведены интенсивные кристаллографические исследования комплексов РНК-полимеразы с другими компонентами системы. Структура полимеразы была установлена сначала с разрешением 2.8 A и затем 2.3 A. Все события развиваются в глубокой щели молекулы полимеразы. Именно сюда попадает дсДНК (двуспиральная ДНК), и здесь локализован активный центр фермента (рис.2). Щель активного центра пересекает, подобно мосту, a-спираль (“мостиковая” спираль) одной из двух больших субъединиц полимеразы.Инициация синтеза РНК состоит из ряда этапов, в которых участвуют общие факторы транскрипции. TBP (TATA-binding protein) сгибает ДНК вокруг С-концевого домена TFIIB, в результате чего образуется комплекс этого белка с полимеразой, и ее активный центр ориентируется так, что оказывается напротив стартового участка транскрипции на ДНК. Далее к комплексу инициации последовательно присоединяются другие транскрипционные факторы - TFIIH и TFIIE. Первый из них расплетает участок ДНК, находящийся в активном центре фермента, а второй удерживает некодирующую цепь расплетенной петли ДНК. При этом ось гибридной ДНК-РНК спирали повернута приблизительно на 90о по отношению к дуплексу ДНК (см. рис.2), вступающему в реакцию транскрипции, благодаря полипептидной “стенке”, которая препятствует прямому прохождению нуклеиновой кислоты через щель. В инициации транскрипции своеобразную роль играет один из доменов TFIIB, названный В-пальцем: он занимает в щели активного центра полимеразы то же место, что и гибрид ДНК-РНК, конкурируя с последним. В начале инициации транскрипции TFIIB стабилизирует гибридный участок, содержащий короткий фрагмент - до пяти нуклеотидов - вновь синтезированной РНК. При дальнейшем увеличении гибридной спирали начинается конкуренция между растущей цепью РНК и TFIIB за пространство в ложбине активного центра полимеразы. По терминологии Корнберга, если побеждает РНК, ее синтез продолжается, и инициация переходит в элонгацию транскрипции: полимераза перемещается за пределы промотора, двигаясь по ДНК. Все факторы транскрипции, кроме IIF, выходят из комплекса инициации. Если же побеждает в конкуренции с РНК TFIIB, синтез РНК прерывается на уровне девяти нуклеотидов, комплекс распадается и инициация начинается заново. Этот феномен, известный ранее как “абортивная” инициация транскрипции, получил объяснение на молекулярном уровне только после исследований Корнберга.
Показаны спираль ДНК (синяя), растущая цепь РНК (красная), ион металла в активном центре в виде фиолетовой сферы и “мостиковая” a-спираль (зеленая).
Кристаллографическое исследование комплексов полимеразы с нуклеотидами позволило описать поэтапно ход присоединения нуклеотида к растущей цепи (рис.3). На первом этапе NTP (нуклеозидтрифосфат) приближается к растущей цепи и связывается в Е-сайте ниже активного центра в обращенной ориентации. Далее происходит вращение нуклеотида, в результате чего он перемещается в А-сайт. В случае комплементарности входящего нуклеотида матричному основанию ДНК на третьем этапе образуется фосфодиэфирная связь. Все завершается транслокацией, необходимой для повторения цикла. Отбор комплементарного нуклеотида контролируется свойствами РНК-ДНК гибрида. Если нуклеотид некомплементарен, он не удерживается в активном центре, и селекция продолжается. Таким образом, в серии кристаллографических исследований комплексов РНК-полимеразы с компонентами системы была установлена на атомном уровне динамическая картина инициации транскрипции.
РНК-полимераза II и медиатор взаимодействуют между собой за счет множества контактов, посредством которых осуществляется перенос сигналов от активатора, связанного с энхансером, к комплексу инициации транскрипции. Начало детальным исследованиям комплекса медиатора с полимеразой положила публикация (незадолго до присуждения Корнбергу премии), в которой рассматривается взаимодействие головной части медиатора из семи субъединиц с ферментом в присутствии фактора транскрипции IIF. Для выяснения динамических взаимодействий на атомном уровне в дальнейшем необходимо будет изучать комплексы разных частей медиатора с компонентами системы инициации транскрипции.
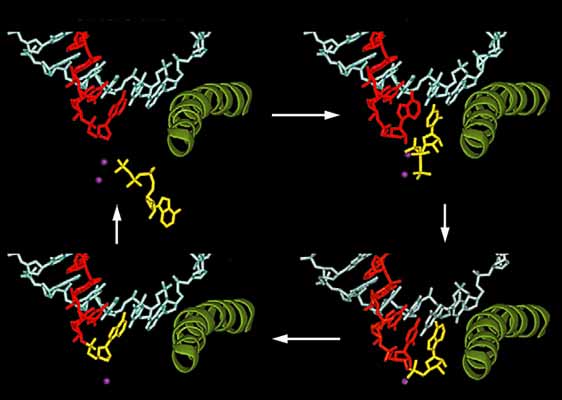
Рис. 3. Четыре кристаллические структуры комплексов РНК-полимеразы II с нуклеозидтрифосфатами.
Изображена область активного центра фермента, в которой находятся транскрибируемая ДНК (синяя), синтезируемый участок РНК (красный), присоединяемый нуклеотид (желтый), ионы Mg (фиолетовые) и пептидная спираль (зеленая). Показаны четыре этапа присоединения нуклеотида к растущей цепи РНК: связывание нуклеотида в Е-сайте (вверху слева); ротация комплементарного нуклеотида в А-сайте (вверху справа); образование фосфодиэфирной связи (внизу справа) и транслокация нуклеотида.Краткое резюме основных достижений Корнберга сводится к следующему. Им установлена общая картина действующего комплекса инициации транскрипции с участием РНК-полимеразы II, общих факторов транскрипции и медиатора, открытого Корнбергом. Промотор за счет ремоделирования хроматина освобождается от нуклеосомы и взаимодействует с ферментом и остальными элементами системы, в результате чего и образуется комплекс инициации транскрипции, состоящий примерно из 60 белков (более 3МДа). Медиатор осуществляет перенос как положительных, так и отрицательных сигналов к транскрипционной машине. Благодаря рентгеноструктурному анализу нынешнему нобелевскому лауреату удалось построить - с атомным разрешением - динамическую картину событий в активном центре РНК-полимеразы II с участием других компонентов системы. В результате было установлено, каким образом осуществляется узнавание промотора, селекция нуклеотидов, образование фосфодиэфирной связи, рост цепи РНК и переход от стадии инициации транскрипции к элонгации.По словам нового нобелевского лауреата, его исследования были бы немыслимы без его сотрудников и помощников, которые могли бы разделить с ним эту высшую научную премию.
© Поляновский О.Л.,
доктор биологических. наук
Институт молекулярной биологии им.В.А.Энгельгардта РАН