№3, 2002 г.
№3, 2002 г.
№3, 2002 г.
№3, 2002 г.
© Н.Б. Рубцов, П.М. БородинЭволюция хромосом: от А до В и обратно
Н.Б. Рубцов, П.М. Бородин
Николай Борисович Рубцов, д.б.н.,
зав. лаб. Института цитологии и генетики СО РАН.Павел Михайлович Бородин, д.б.н,
зав. секцией рекомбинационного и сегрегационного анализа того же института,
профессор кафедры цитологии и генетики НГУ.Что такое хромосомы, сейчас, наверное, знают уже все. По крайней мере в школьном учебнике про них написано довольно подробно. Набор хромосом (кариотип) - надежная характеристика видовой принадлежности животных и растений. У всех представителей каждого вида их количество одинаково. Это верно для большинства видов, но далеко не для всех: у многих животных и растений наряду с хромосомами основного набора (А-хромосомами) обнаруживаются добавочные, или сверхчисленные хромосомы, так называемые В-хромосомы. Их число, форма и размер могут быть разными у представителей одного и того же вида (у одних - очень много, а у других - не быть вовсе). Так, у серебристо-черных лисиц количество В-хромосом варьирует от 0 до 6, а у азиатской лесной мыши (именно об этом виде - Apodemus peninsulae - и пойдет дальше речь) - от 0 до 17. Сейчас известно около 500 видов животных с В-хромосомами. В кариотипах млекопитающих В-хромосомы обнаружены у многих видов грызунов, некоторых хищников и копытных. У человека, как и у остальных приматов, В-хромосом нет. Наличие или отсутствие добавочных хромосом, как правило, никак не сказывается на фенотипе - внешнем виде и других свойствах животного.
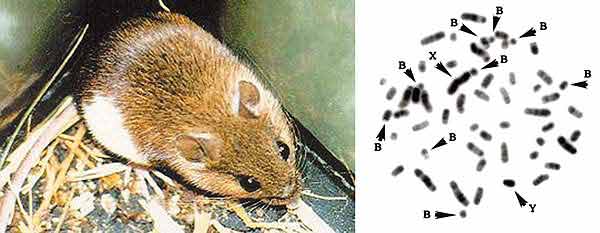
Этот факт поначалу очень смущал исследователей, убежденных, что геном есть чертеж организма и поэтому устроен мудро, экономно и в нем нет ничего лишнего. Исходя из этого убеждения, генетики старались найти, чем В-хромосомы могут быть полезны их носителям. Такие поиски редко заканчивались успехом, и вопрос о том, для чего нужны В-хромосомы, оставался без ответа.Азиатская лесная мышь (слева) и ее кариотип,
который содержит 23 пары аутосом, половые хромосомы Х и Y и 9 В-хромосом.Применение методов молекулярной генетики к анализу А- и В-хромосом показало, что и те и другие содержат огромное количество многократно повторенных и, видимо, не кодирующих белки последовательностей ДНК. Вся разница между ними состоит в том, что в А-хромосомах наряду с некодирующими повторами довольно много функциональных генов (хотя и не так много, как думали), а в В-хромосомах они вовсе отсутствуют или их число ничтожно мало. На этом фоне вопрос, зачем нужны В-хромосомы, потерял свою остроту. Прежде всего следовало понять, почему в геноме так много бессмысленных повторенных последовательностей, и нет ли в этом изобилии некоего глубокого, но скрытого от нас смысла?
История о том, как искали ответ на этот вопрос, чрезвычайно интересна и драматична. Однако ее изложение увело бы нас далеко от В-хромосом. Поиски ответа привели к заключению, что никакого смысла в бессмысленной ДНК нет. Убеждение, что геном устроен мудро, экономно и на благо организма, рухнуло. На смену ему пришла концепция эгоистического гена, в рамках которой повторенные последовательности рассматриваются как геномные паразиты, использующие клеточную машинерию для собственного размножения. Вначале концепция была чисто умозрительной, не более чем красивой метафорой, которая избавляла от необходимости поисков функционального значения избыточной ДНК. Вскоре, однако, выяснилось, что метафора оказалась довольно точной.
Было обнаружено, что большая часть бессмысленных последовательностей составлена из паразитических, вирусоподобных элементов, которые могут перемещаться по геному и размножаться как в составе хромосомной ДНК, так и вне ее. Такие мобильные генетические элементы используют механизмы генетической рекомбинации для перемещения по геному и размножения.
Рекомбинация - это процесс перетасовки генов между родственными хромосомами или внутри хромосомы. Гены, вовлеченные в этот процесс, должны быть хотя бы частично гомологичны друг другу, т.е. иметь весьма похожие участки, которые опознают друг друга и сближаются. В местах контакта возникают разрывы в молекулах ДНК и между ними происходят обмены. Затем разрывы залечиваются и целостность хромосом восстанавливается.
Мобильные элементы находят в ДНК хозяина короткие участки, гомологичные собственным последовательностям, и лавинообразно встраиваются в его геном. Чем больше их копий присутствует в геноме, тем больше число паразитических последовательностей и, значит, выше шанс новых перемещений. В ходе своих блужданий по геному, мобильные элементы могут прихватывать куски хозяйских генов и перетаскивать их с места на место.
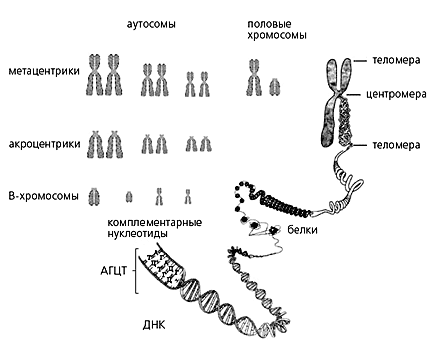
Организация хромосом млекопитающих. Хромосома представляет собой комплекс ДНК и белков. ДНК содержит генетическую информацию, необходимую для создания и функционирования организма, а белки упаковывают ДНК, причем неравномерно - одни районы плотнее других. Плотность упаковки меняется в ходе клеточного цикла. Перед началом клеточного деления хромосомы компактизуются. Каждая хромосома содержит центромеру - особый участок ДНК, к которому прикрепляются специфические белки, а к ним в свою очередь нити веретена клеточного деления. Концы хромосомы защищены теломерами. Между центромерой и теломерой находится плечо хромосомы. Положение центромеры определяет тип хромосомы. Хромосомы с центромерой на конце называются акроцентрическими (или актроцентриками), а те, у которых центромера где-то посредине - метацентрическими (или метацентриками).Выяснилось, что А-хромосомы животных и растений насыщены паразитическими элементами, а В-хромосомы практически целиком состоят из них. Поэтому В-хромосомы можно рассматривать как шайку геномных паразитов. В таком случае вопрос о том, для чего нужны В-хромосомы, теряет актуальность: паразиты они и есть паразиты, и с этим ничего не поделаешь. Хозяева пытаются избавиться от паразитов, которые, как правило, сопротивляются. Начинается долгая и безнадежная гонка вооружений между паразитами и хозяевами, ускоряющая эволюцию и тех, и других, но не приносящая никому ни победы, ни счастья. Хозяева совершенствуют замки от паразитов, а паразиты находят к ним отмычки. В том, что мобильные элементы используют для своего размножения генетическую рекомбинацию, есть определенная доля злой шутки. Ведь основное предназначение рекомбинации в том и состоит, чтобы, перетасовывая гены, постоянно менять шифр на кодовых замках клетки. И именно ее паразиты используют как отмычку. Но это, как пишут в романах, отдельная история.Вернемся к нашим В-хромосомам. Назвав их паразитами, мы избавили себя от необходимости отвечать на вопрос, зачем они нужны. Но понять, откуда они берутся и как собственно паразитируют на А-хромосомах, по-прежнему важно.
Пытаясь ответить на эти вопросы, мы выбрали в качестве модели В-хромосомы азиатской лесной мыши, чтобы выяснить, есть ли у последовательностей ДНК В-хромосом родственники в А-хромосомах.
Родство между молекулами ДНК определить довольно просто. Как известно, ее двойная спираль составлена из двух цепей нуклеотидов А, Т, Г и Ц, которые зеркально отражают друг друга. Напротив А всегда стоит Т, напротив Г стоит Ц, и наоборот. Поэтому по одной цепочке всегда можно сделать вторую. Такие “зеркальные” или, как их называют, комплементарные участки ДНК удерживаются вместе связями между А-Т и Г-Ц. При высокой температуре нити ДНК расходятся, а при ее понижении связи восстанавливаются и спирали объединяются вновь. При этом образование новых двойных спиралей прямо пропорционально комплементарности цепей. Это свойство ДНК используется в методе гибридизации нуклеиновых кислот in vitro - на стекле. Не вдаваясь в теорию и технические сложности метода, расскажем о наших экспериментах.
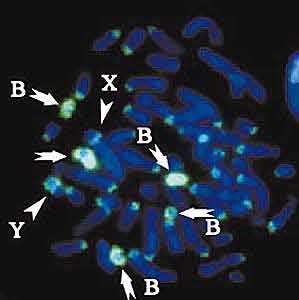
Клетки полевой мыши наносили на предметное стекло. Сверхтонкой стеклянной иглой соскребали со стекла все В-хромосомы одной клетки. Каждую хромосому помещали в отдельную микрокаплю, объем которой был в 2 тыс. раз меньше дождевой капли. Затем из каждой хромосомы выделяли ДНК и размножали (амплифицировали) в пробирке. При этом использовали нуклеотиды, к каждому из которых присоединяли молекулы флуоресцентного красителя. Такую меченую ДНК, выделенную из конкретной хромосомы или отдельного ее района, называют ДНК-пробой. Итак, получали набор, который включал пробы для каждой отдельной В-хромосомы и для прицентромерных участков А-хромосом. Кроме того, сделали три пробы, одна из которых содержала ДНК хромосомы В2, вторая - одного ее плеча, а третья - другого, но ни одна из них не включала материала из центромерной области этой хромосомы. Разные пробы метили красителями разного цвета: например, проба на центромерные районы А-хромосом была красной, а на В-хромосомы - зеленой.
ДНК этих проб соединяли с комплeментарными участками расплетенной ДНК хромосом, распластанных на предметном стекле, и анализировали под микроскопом. Флуоресцентный сигнал наблюдался в тех районах хромосом, где меченая красителем ДНК сплелась с идентичными или родственными ей копиями ДНК хромосом.
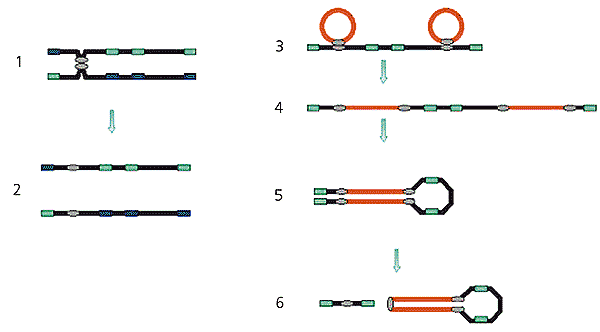
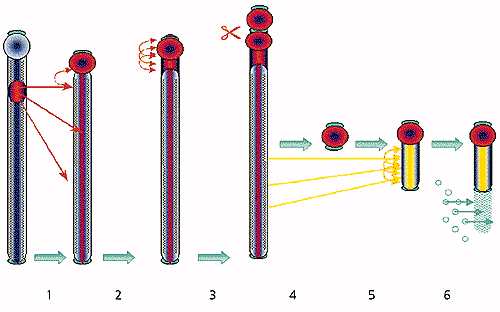
Рекомбинация между молекулами ДНК, имеющими гомологичные (сходные) последовательности (серый овал). Две молекулы сближаются (1) и обмениваются участками (2). Мобильные генетические элементы (красные линии), которые содержат последовательности, гомологичные последовательностям хромосомной ДНК, могут с ней спариваться (3). В результате рекомбинации мобильные элементы встраиваются в хромосомную ДНК (4). Повторенные участки мобильных элементов гомологичны друг другу, поэтому они могут спариваться, рекомбинировать (5) и покидать хромосому, включая в свой состав участки хромосомной ДНК (6).Кроме того, почти все В-хромосомные пробы интенсивно окрашивали прицентромерные области А-аутосом (всех А-хромосом, за исключением половых); и в половых хромосомах - районы плотной упаковки ДНК, которые находились на конце Y-хромосомы и внутри плеча Х-хромосомы и содержали большое количество повторенных последовательностей ДНК. В то же время проба, сделанная из ДНК центромерных районов А-хромосом, окрашивала прицентромерные районы крупных В-хромосом, почти целиком мелкие В-хромосомы и те же самые районы на половых хромосомах, которые окрашивались пробами, сделанными из В-хромосом. Это означает, что в прицентромерных районах В-хромосом имеются повторы, родственные последовательностям, находящимся в прицентромерных районах аутосом и в плотных блоках на половых хромосомах.
Но этим родство между А- и В-хромосомами не ограничивается. Когда на препарат наносили пробы В-хромосом, цветную метку обнаруживали не только в прицентромерных районах аутосом, но и на их плечах, хотя в А-хромосомах они окрашивались во много раз слабее, чем в В-хромосомах. Особенно показательными оказались эксперименты с пробами, которые содержали ДНК плеч хромосомы В2: они сильно окрашивали плечи этой и других В-хромосом и очень слабо - плечи аутосом, в то время как центромерные районы А- и В-хромосом оставались неокрашенными. Значит, в плечах А-хромосом в небольшом количестве содержатся повторы ДНК, составляющие основную массу ДНК плеч В-хромосом.
Кстати, использование таких специфических проб позволило выявить одну интересную особенность строения В-хромосом. Оказалось, что проба из одного плеча хромосомы одинаково интенсивно окрашивает оба плеча. Такие хромосомы с идентичными плечами называются изохромосомами. Они иногда возникают в результате хромосомных перестроек в раковых клетках, в клетках, культивируемых in vitro, но почти никогда не встречаются в норме. Что же касается В-хромосом нашей лесной мыши, то, похоже, почти все они - изохромосомы: имели плечи равных размеров, которые симметрично окрашивались любыми пробами. Исключение составляли, может быть, только самые мелкие хромосомы, в которых почти невозможно разглядеть детали морфологии.
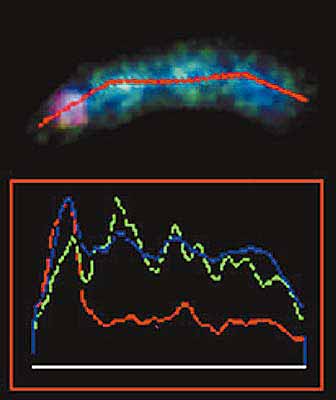
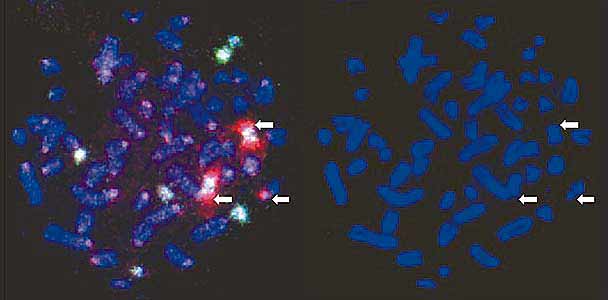
Две такие мелкие В-хромосомы преподнесли нам занятный сюрприз. Когда препарат окрашивали ДАПИ - красителем, выявляющим любую ДНК безотносительно к ее последовательности, - они выглядели, как точечки. Казалось, что эти хромосомы состоят только из центромер и вовсе не имеют плеч. Однако, когда на препарат нанесли сделанные из них пробы, специфически окрашивающие их ДНК, обнаружили, что одна окрашивает тела хромосом, а другая выявляет красивые нимбы (вообще-то они больше похоже на метелки, но “красивая метла” как-то не звучит, а “нимб” - звучит) вокруг родной хромосомы и около концов еще двух В-хромосом. Причем нимбы распространялись далеко за пределы концов хромосом, которые мы в специальном эксперименте пометили пробами на теломерные последовательности ДНК.
Пробы ДНК, приготовленные из мелких В-хромосом, были мечены красным и зеленым цветами. Одна из них (зеленая) нормально окрашивала тела В-хромосом. Вторая (красная) очень странно окрашивала родную хромосому и концы двух других хромосом. Она выявляла нити ДНК, простиравшиеся далеко за физические концы хромосом. В этом можно убедиться, сравнив В-хромосомы, окрашенные только ДАПИ (справа), и те же хромосомы, окрашенные ДАПИ и ДНК-пробами (стрелки, указывающие на В-хромосомы, находятся в идентичных позициях).Для того чтобы понять природу нимбов, необходимо сделать небольшой экскурс в проблему упаковки хромосом. Мы уже говорили о том, что хромосомы представляют собой комплексы ДНК с белками и плотность упаковки ДНК зависит от ее функциональной активности. В момент деления клеток активность генов прекращается и ДНК уплотняется. Белки, присоединяясь к определенным последовательностям ДНК, приклеивают ее петли к белкам хромосомного скелета, а затем склеивают образовавшиеся при этом завитки. В таком плотно упакованном виде хромосома легко переносит манипуляции клеточного деления на две равные части и расхождение их к разным полюсам клетки. Можно думать, что ДНК нимбов не содержит последовательностей, необходимых для присоединения упаковочных белков, и остается в расплетенном состоянии, когда вся остальная ДНК уплотняется. В таком виде она легко доступна для гибридизации с ДНК пробы и ярко окрашивается. А поскольку концентрация ДНК и белков хромосомы на единицу площади предметного стекла в районе нимбов ниже, чем в нормально упакованных хромосомах, ДАПИ окрашивает нимбы настолько слабо, что увидеть эту окраску не удается.Кроме концов некоторых В-хромосом такая проба окрашивала два небольших симметрично расположенных участка в двух плечах одной из В-хромосом. Тогда мы наблюдали не нимб, а интенсивное локальное окрашивание (почему это так, мы обсудим чуть позже). Никаких следов связывания ДНК этой пробы с ДНК А-хромосом мы не обнаружили.
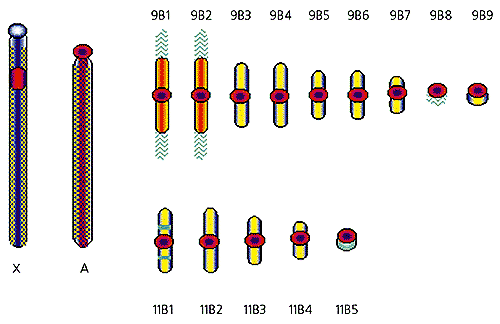
Итак, результаты окрашивания хромосом лесной мыши В-хромосомными пробами показывают, что в геноме этого вида присутствуют по крайней мере три класса повторов:
Все три класса повторов встречаются в В-хромосомах не только у тех животных, из которых получали пробы, но и у других изученных нами лесных мышей Apodemus peninsulae. Следовательно, они общие для данного вида, а может, и всего рода Apodemus.
- один локализован в прицентромерных районах аутосом и В-хромосом, а так же в двух районах половых хромосом;
- другой составляет основную массу плеч В-хромосом, а в А-хромосомах содержится в значительно меньших количествах;
- третий обнаруживается на концах некоторых В-хромосом и ведет себя странно, образуя нимбы (или метелки, если этот образ вам больше нравится).
Чтобы в этом убедиться, мы гибридизовали наши пробы ДНК Apodemus peninsulae с ДНК хромосом полевых мышей родственного вида Apodemus agrarius, у которых В-хромосомы встречаются крайне редко. Из тысяч исследованных полевых мышей только у двух нашли В-хромосомы - да и то всего по одной. У той мыши, от которой брали клетки для анализа, В-хромосом не было вовсе. Мы уже знали, что прицентромерные повторы В-хромосом у Apodemus peninsulae имеются и в В-, и в А-хромосомах, и поэтому мы ожидали их увидеть и в А-хромосомах Apodemus agrarius. К сожалению или к счастью, наши ожидания не оправдались. У этого вида центромерные районы не окрашивались даже в том случае, когда использовали пробу на центромерные районы А-хромосом Apodemus peninsulae. И эта проба, и пробы В-хромосом слабо и дисперсно окрашивали только плечи А-хромосом Apodemus agrarius. Единственными местами, где мы видели более или менее яркое окрашивание, были плотные блоки на половых хромосомах. Они, как вы помните, ярко окрашивались этими пробами и у Apodemus peninsulae.
Три типа повторенных последовательностей: прицентромерный (красный), типичный для плеч В-хромосом (желтый) и типичный для концов некоторых В-хромосом (зеленый).
Поскольку такие блоки есть у обоих видов, можно предположить, что они унаследованы от общего предка, который, как показали специальные исследования, существовал 8-7 млн лет назад. Не в них ли скрывались до поры до времени мобильные паразитические последовательности ДНК, которые потом образовали В-хромосомы Apodemus peninsulae?Попытаемся представить себе, как это было. Из половых хромосом мобильные элементы проникли в аутосомы обоих видов. Для этого им нужно было найти в ДНК хозяина участки, сходные с их собственными последовательностями. У Apodemus agrarius такие участки нашлись в плечах аутосом, а у Apodemus peninsulae - в прицентромерных районах. Поскольку эти районы всегда содержат множество повторов, можно думать, что тех, которые имели гомологию с мобильными элементами, было немало. Это сильно облегчило внедрение и последующее размножение паразитических последовательностей. Есть еще одна особенность прицентромерных районов, облегчающая жизнь паразитов. Дело в том, что там, как правило, нет активных генов, и потому внедрение паразитов не нарушает функции генома. Совсем иная ситуация в плечах хромосом, где находятся гены, отвечающие за синтез белков. Внедрение паразита внутрь гена или даже рядом с ним может привести к непоправимым нарушениям жизненно важных функции и к гибели клеток или организмов, в которых такое событие произошло.
Итак, в геноме Apodemus peninsulae паразитическим мобильным элементам сильно повезло. Они проникли в прицентромерные последовательности и в ходе своего размножения захватили специфические последовательности хозяйской ДНК - центромерные и теломерные. Это открыло дорогу для возникновения В-хромосом. Ведь что нужно фрагменту ДНК, чтобы стать хромосомой? Иметь центромеру, дабы на равных с нормальными хромосомами участвовать в клеточном делении, и две теломеры, дабы защитить свои концы от разрушения. Некоторые из В-хромосом Apodemus peninsulae (самые мелкие) состоят в основном из центромеры с прицентромерными повторами и пары теломер - и очень хорошо себя чувствуют. Это - или самые молодые из В-хромосом, или реликты ранних этапов хромосомной эволюции.
Как только В-хромосомы возникли в такой примитивной форме, они стали “землей обетованной”, куда устремились и другие мобильные паразитические элементы, влачившие до той поры жалкое существование в составе плеч аутосом или за пределами хромосомной ДНК. Последовательности, которые составляют основную массу ДНК плеч В-хромосом Apodemus peninsulae, присутствуют в небольших количествах и в плечах аутосом обоих видов мышей. Но там их размножение сдерживается естественным отбором. Иное дело в В-хромосомах - здесь им живется и размножается гораздо вольготнее. У большинства В-хромосом прицентромерные и плечевые последовательности ДНК четко разграничены. Однако в некоторых В-хромосомах (самых старых или самых эволюционно продвинутых?) они оказались перемешанными.
Последними в В-хромосомы пришли концевые повторы, те самые, что образуют метелки. Именно эта способность, а вернее, неспособность плотно упаковываться в момент клеточного деления выдает их экстрахромосомное происхождение. По-видимому, они еще не успели включить в свой состав те последовательности хромосомной ДНК, с которыми связываются белки компактизации. Как вы помните, тот же класс повторов мы обнаружили внутри плеч одной из В-хромосом, где он был нормально упакован. Может быть именно потому он и был упакован, что находился внутри, а не на конце хромосомы. Белки склеивали и соседние с ним районы, и его заодно. А может быть, он попал в середину хромосомы уже давно и успел за счет рекомбинации приобрести последовательности, необходимые для связывания с упаковочными белками.
Таким образом, вполне вероятно, что В-хромосомы, которые мы нашли в геноме лесной мыши, возникли не одновременно. Одни появились раньше, другие позже, да и сами пути их эволюции были разными. Можно ли выяснить, когда это произошло?
До сих пор мы рассматривали только два вида рода Apodemus. Но в нем много видов, и В-хромосомы часто встречаются не только у Apodemus peninsulae. Они обнаруживаются и во многих (хотя и не во всех) популяциях европейской лесной мыши Apodemus flavicollis. Не значит ли это, что оба вида унаследовали В-хромосомы от общего предка? Нет, не значит. Дело в том, что Apodemus peninsulae и Apodemus flavicollis принадлежат к разным подродам, которые разделились 10-8 млн лет назад. Внутри каждого подрода большинство видов не имеют В-хромосом вовсе. Да и популяции Apodemus peninsulae и Apodemus flavicollis и особи внутри этих популяций очень сильно отличаются друг от друга по числу и форме В-хромосом. Логичнее предположить, что эти виды унаследовали от общего предка не сами В-хромосомы, а способность создавать их de novo и толерантность к их присутствию.
Работа выполнена при поддержке Российского фонда фундаментальных исследований и программы Университеты России. Проекты 99-04-49960, 01-04-48875, 01-04-49534 и 015.07.01.024.