№ 3, 2003 г.
© Ю.А. Лабас, А.В. Гордеева, А.Ф. Фрадков
Флуоресцирующие и цветные белки
Ю.А. Лабас, А.В. Гордеева, А.Ф. Фрадков
Юлий Александрович Лабас, кандидат биологических
наук,
ведущий научный сотрудник Института биохимии им.А.Н.Баха
РАН.
Анна Викторовна Гордеева, аспирантка того же института.
Аркадий Федорович Фрадков, кандидат химических
наук, старший научный сотрудник
Института биоорганической химии им.М.М.Шемякина и Ю.А.Овчинникова
РАН.
По приказу короля 4 января 1761 г. датский военный корабль направился из Копенгагена в Аравию с научной экспедицией. В ее составе были трое ученых, в их числе зоолог П.Форскал, а кроме того - художник, врач и слуга. Однажды, в начале марта, когда корабль плыл еще по Северному морю, пассажиры заметили в воде странное свечение. Причиной оказались небольшие, с крупную монету величиной, очень прозрачные медузы. По ночам при волнении воды они ярко светились зеленым фосфорическим светом. Форскал заспиртовал несколько экземпляров медуз и записал по-латыни в походном дневнике, что при раздражении и гибели они светятся. С этой записи началась история исследований Medusa aequorea, как назвал ее Форскал. Под этим именем она вошла и в его посмертно изданную монографию “Fauna arabica” (1775). В 1809 г. в названии появилась фамилия первооткрывателя - Aequоrea forskalea. В новейших индексах цитат и в Интернете значится ее последнее устоявшееся имя - Aequorea victoria.
Оказалось, что у медузы имеется светящееся вещество. Через полтора века выяснилось: для предварительной “заправки” эквореи таким веществом абсолютно необходим кислород. Само же оно представляет собой целентеразин (от лат. Coelenterata - кишечнополостные) - имидазолпиразиновое производное.

Светящаяся в темноте медуза Aequorea victoria [12].
Целентеразин окисляется молекулярным кислородом до гидроперекиси в момент присоединения к экворину, белку, способному связывать ионы кальция. C конца 60-х годов экворин обрел всемирную известность как сверхчувствительный индикатор этих ионов внутри клетки. Впрочем, с 1993 г. куда популярнее стал зеленый флуоресцирующий белок (green fluorescent protein - GFP), тоже впервые выделенный из A.victoria. Разговор далее пойдет об этих и близких к ним белках.
Зеленый подарок эквореи
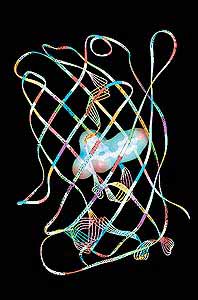
В 1962 г. О.Шимомура, Ф.Джонсон и Ю.Сайга отметили разницу между цветом биолюминесценции живой эквореи и выделенного из нее комплекса светящегося вещества с экворином [1]. У живой медузы свечение зеленое с максимумом 508 нм. А чистый комплекс, реагируя с ионами кальция, испускает синий свет (максимум 465 нм). Оказалось, в светящихся энтодермальных клетках эквореи присутствует, кроме экворина, еще и другой - зеленый флуоресцирующий белок. Если облучить его синим или ближним ультрафиолетовым светом (в спектре возбуждения максимум 395 нм, второй пик - 475 нм), он излучает зеленый свет с максимумом 508 нм. Исследователи установили аминокислотную последовательность зеленого белка (его молекулярная масса составляет 28 кДа) и выяснили, что он состоит из 238 остатков. Квантовый выход флуоресценции GFP фантастически велик: около 0.8! Следует отметить, что энергия возбуждения экворина отчачти мигрирует на GFP безызлучательно благодаря непосредственному контакту с двумя молекулами GFP (он и существует в виде димера при одной молекуле эквoрина).
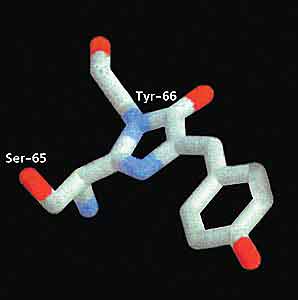
В 1992 г. американские ученые клонировали ген зеленого флуоресцирующего белка и внедрили его в геном кишечной палочки (Esherichia coli) [2]. В результате был получен белок в количестве, нужном для его исследований, в том числе для расшифровки аминокислотной последовательности, и выявлены совершенно уникальные особенности GFP. Оказалось, он, в отличие от всех до того известных окрашенных и флуоресцирующих белков (хромопротеидов: фикобиллинов, фикоэритринов, каротенопротеинов), для обретения оптических свойств не нуждается в какой-либо внешней хромофорной группе или в солях тяжелых металлов. Извне для его “созревания” - появления яркой зеленой флуоресценции - требуется только молекулярный кислород. Этот процесс длится довольно долго. Детектируемая флуоресценция появляется через полтора часа после синтеза, а достигает максимума только через несколько часов. При “созревании” в пептидной цепи белка образуется р-гидроксибензилиден-имидазолиноновое кольцо из трех аминокислотных остатков, следующих друг за другом: серина-65, тирозина-66 и глицина-67. Оно-то и служит хромофорной группой.
В ходе такой реакции Тир-66 автокаталитически дегидрируется (отдает два протона) и окисляется молекулярным кислородом. Не вошедшая в состав хромофора часть пептидной цепи белка скручивается в подобие бочки или клетки, внутри которой помещается хромофор. Стенки “бочки” образованы 11 b-складками, а крышка, дно и связки с хромофором - a-спиральными участками белковой молекулы.
Кроме зеленого белка эквореи, подобные ему существуют у десятков видов других кишечнополостных животных: колониальных гидроидных (Obelia longissima, O.geniculata, Clytia sp. и др.) и коралловых полипов - морских перьев (Renilla muelleri, R.reniformis, Ptilosarcus sp., Stylatula elonga, Acanthophtilum gracile и т.д.). У всех перечисленных организмов синий цвет преобразуется в зеленый.
Между прочим, у этих бесскелетных кораллов механизм импульсной биолюминесценции не такой, как у гидроидов. Люциферин (целентеразин) у них светится в комплексе не с белком, связывающим ионы кальция, а с ферментом люциферазой. С ней-то у коралловых полипов (Anthozoa) и образует комплекс зеленый флуоресцирующий белок (с более высоким сродством, чем у эквореи). Судя по спектрам биолюминесценции и флуоресценции живых фотогенных клеток, он, видимо, есть у организмов, даже не принадлежащих к типу кишечнополостных, например у гребневиков Bolinopsis infundibulum [3].
Охота за разноцветными белками
Осенью 1998 г. С.А.Лукьянов, руководитель исследовательской группы Института биоорганической химии РАН, обратился к одному из авторов этой статьи - Ю.А.Лабасу - с просьбой порекомендовать биолюминесцентный организм, в котором можно найти белок с необычным цветом флуоресценции.
Лабас, обдумывая эту задачу, обратил внимание на следующие обстоятельства. Классы кишечнополостных Hydrozoa и Anthozoa, у которых обнаружены GFP, разошлись еще в глубоком докембрии. Гребневики от общих с ними предков отделились и того ранее. Следовательно, весьма вероятно, что все эти организмы унаследовали GFP от общих небиолюминесцентных предшественников. В то же время крупные подвижные животные с хорошо развитым зрением - рыбы, головоногие моллюски, высшие раки (т.е. потенциальные враги, а значит, и эволюционный повод обретения биолюминесценции) - появились не раньше кембрийского периода.
1 2 3
4 5 6
7 8 Флуоресцирующие и окрашенные морские животные. Из числа первых здесь показаны актиния Anemonia majano с ярко-зеленой флуоресценцией щупалец (1), кораллы Ptilosarcus gurney (2) и Ricordia yuma (3), флуоресцирующие соответственно желтым и красным; из вторых - Acropora humilus (4), Actinia equina (5), Corynactis californica (6), Zoanthus sp. (7) и Anemonia sulcata (8), у которых тело и кончики щупалец окрашены по-разному (№3, 6 - фото А.О.Романько, остальные взяты из Интернета http://www.google.com.ru).Стало быть, у GFP-подобных белков, как и у других компонентов системы свечения (люциферинов, люцифераз и т.д.), могли быть какие-то добиолюминесцентные функции. Почему бы им не сохраниться и по сей день? В самом деле: у целого ряда несветящихся кораллов, живущих в морских аквариумах Московского зоопарка и у любителей, при освещении ультрафиолетом или голубым светом появляется яркая зеленая флуоресценция всего тела или только отдельных участков. На вид она очень похожа на флуоресценцию GFP той же экворины. Разные исследователи полагали, что за флуоресценцию кораллов ответственны какие-то низкомолекулярные солнцезащитные “пигменты”. А вдруг это не так?Ведь у них флуоресценция бывает не только зеленой, но и синей, желтой, оранжевой, рубиново-красной. Более того, она может замещаться обычной окраской. Например, у одного из двух видов актиний, которых часто содержат в морских аквариумах, а именно у Anemonia majano, кончики щупалец ярко флуоресцируют зеленым, а у другого - A.sulcata - просто окрашены в пурпурный цвет. Не связано ли это у анемоний с разными вариантами белковых молекул, близких по структуре GFP?
Сколь ни безумными казались эти мысли, мы начали поиск таких белков. Сначала выделили матричную РНК (мРНК) из ярко окрашенных участков тела шести разных видов мягких кораллов - в первую очередь из флуоресцирующих кончиков щупалец актинии А.majano, жившей в домашнем морском аквариуме известного московского любителя А.О.Романько. Потом сотрудники Лукьянова разнообразными молекулярно-биологическими манипуляциями получили из мРНК кодирующую ДНК (кДНК) и попробовали “выловить” из нее молекулы, сходные по нуклеотидной последовательности с геном зеленого белка эквореи.
“Ловля” осуществлялась с помощью праймеров - синтезированных одноцепочечных кусочков этого гена, способных заякорить на себе комплементарную цепь ДНК. Синтезировали для такой цели участки в 20 - 25 нуклеотидов, интуитивно показавшиеся самыми консервативными в гене GFP A.victoria. Поиски увенчались успехом, после чего ген пересадили в геном кишечной палочки. И она заблистала ярким зеленым цветом!
Вскоре мы узнали от Романько, что в его морском аквариуме живет Ricordia yuma - удивительная дискосома (дисковидная актиния без щупалец), которая при синем освещении флуоресцирует ярким красно-оранжевым цветом (у других дискосом он сине-зеленый). Само собой разумеется, мы заподозрили, что и у этой дискосомы есть белок, подобный зеленому флуоресцирующему. Далее дело пошло быстрее. Из гена GFP A.majano и прочих подобных генов легче было конструировать праймеры для все новых вариантов.
Гены вновь открытых белков и GFP A. victoriа оказались гомологичными: последовательности их нуклеотидов совпадали на 20 - 30%. Значит, такой же гомологией характеризовались и сами белки. Весьма солидная степень совпадения, если учесть, что ветви кораллов и гидроидов разделились примерно полмиллиарда лет назад! Вероятность не поймать праймером фрагмент гена с полным совпадением нуклеотидов при первой попытке М.В.Матца была громадной. Но - чудо. Иначе не скажешь. Одним словом, в самом начале охоты за генами белков, подобных зеленому флуоресцирующему, всем участникам сопутствовало фантастическое везение.
В дальнейшем работа стала почти рутинной. И вот ее сенсационный результат: за большинство флуоресцентных и даже обычных окрасок (а это все цвета радуги!) несветящихся видов кораллов и, возможно, других кишечнополостных ответственны вовсе не разнородные низкомолекулярные “пигменты” и их комплексы с белками, как полагали ранее, а своеобразные белки одного семейства с GFP. У них одинаковые или очень близкие молекулярная масса, число аминокислотных остатков (229 - 266) и, что куда важнее, их последовательность. Та же самая бочкообразная структура и, главное, сходным образом устроенный хромофор с участием Тир-66 и Гли-67. Только в 65-й позиции вместо серина, как у A.victoria, находятся другие аминокислоты: лизин, аспарагин или глютамин.
Но почему же тогда у новых белков цвет флуоресценции бывает не только зеленым, но и желтым, красным и т.д.? Почему некоторые из них вообще не флуоресцируют, а просто ярко окрашивают животных в малиновый, синий, фиолетовый цвета? Выяснилось: это зависит главным образом от аминокислотных остатков, расположенных не в хромофоре, а за его пределами - в бочковидной структуре молекулы. Оптические свойства такой конструкции очень трудно понять и тем более предсказать. К тому же оказалось, что сам хромофор в зеленых, желтых и красных флуоресцирующих белках имеет разную структуру, хотя в его состав входят те же тирозин и глицин.
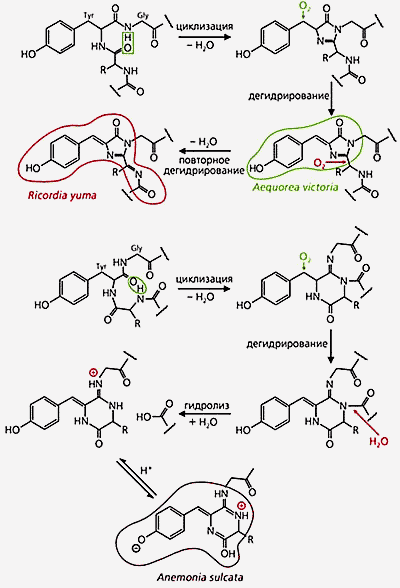
Формирование хромофорных групп в молекулах двух флуоресцирующих белков (вверху) и одного цветного белка [14]. Образование тех и других хромофоров начинается с циклизации, обусловленной отщеплением молекулы воды от соседних аминокислот. Последующее дегидрирование приводит к возникновению хромофора зеленого флуоресцирующего белка у эквореи (Aequorea victoria), а если дегидрирование повторяется, образуется хромофор, который обеспечивает красную флуоресценцию у дискосомы (Ricordia yuma). Малиновый цвет щупалец у Anemonia sulcata обусловлен хромофором, процесс образования которого сложнее: после циклизации и дегидрирования происходят гидролиз и последующее присоединение атома водорода. На схемах обозначены две аминокислоты, входящие в состав хромофора, - тирозин (Tyr) и глицин (Gly).К настоящему времени коллективом исследователей Лукьянова и Матцем (биостанция Витней, Флорида, США) клонировано уже более 30 генов GFP-подобных белков из разных видов несветящихся кораллов [4]. Получены десятки мутантов. Большинство этих белков ответственно за цветную (синеватую или зеленую, желтую, оранжевую, красную) флуоресценцию всего животного или отдельных участков тела. Другие (их пока девять) - за пурпурную или голубую окраску. Это могло быть связано с наличием по соседству с хромофором аминокислотных остатков, содержащих SH-группы (т.е. цистеин или метионин). Однако, судя по флуоресцирующим мутантам, причина окрашивания не в них.С цветными белками еще далеко не все ясно. Некоторые из них сами не флуоресцируют, например пурпурный белок из A.sulcata, а просто окрашивают животное. Но стоит посветить на них зеленым светом, и разгорается яркая красная флуоресценция. Синий свет ее немедленно гасит. Природа таких странных явлений пока не объяснена.
Получены мутанты, которые после разовой лазерной засветки так и остаются флуоресцентными. Этими белками-маркерами можно метить цитоскелетные и другие белки, чтобы наблюдать за их перемещениями в клетке. Удивительное свойство всех GFP-подобных белков - их поразительная устойчивость к изменениям температуры, кислотности среды (рН), к разрушающим белки ферментам протеазам и разного рода химическим соединениям, которые вызывают денатурацию белков.
Не на Барьерном рифе сыскали мы наш живой материал, а у московских любителей экзотической фауны из Индийского и Тихого океанов, в Московском зоопарке и московских зоомагазинах. А ведь годом позже выяснилось, что нам дышала в затылок группа австралийских ученых, которые работали с объектами, добытыми из моря. Австралийцы, однако, шли совсем другим путем [5]. Выделили сначала некий голубой нефлуоресцирующий белок из мадрепоровых кораллов и тщетно пытались отделить от него хромофорную группу. А тут заметили, что молекулярная масса у их белка, почти как у GFP. Начали смотреть аминокислотную последовательность с N-конца белка. Видят: опять отдаленно напоминает GFP. И тогда они догадались с чем имеют дело. В 2000 г. в Германии тоже были выделены цветные белки из актинии A.sulcata, и они проявляли черты сходства с GFP [6]. Задержись наше первое сообщение в “Nature Biotechnology” [7], и мы потеряли бы приоритет.
Зачем кораллам яркая окраска?
Как ни странно, вразумительный ответ на этот вопрос по сей день отсутствует. Довольно обычные цвета нефлуоресцентной окраски кораллов - голубой, зеленый, желтый или бурый. Гораздо чаще - ярко-красный, глубже 15 - 30 м кораллы бывают почти серыми. Флуоресцируют же по большей части зеленым или оранжево-красным. Цветные и флуоресцирующие белки сосредоточены у кораллов в особых гранулах внутри клеток. Группа австралийских ученых, возглавляемая бывшей москвичкой А.Селих, отметила, что при ярком солнечном освещении такие гранулы в двухслойном теле кораллов располагаются над внутриклеточными симбиотическими водорослями зооксантеллами. В слабоосвещенных зонах аналогичные гранулы находятся в основном под водорослевыми клетками. Поэтому Селих и ее коллеги утверждают, что цветные белки служат своеобразным экраном, который предотвращает чрезмерную активацию фотосинтеза у зооксантелл при слишком ярком свете [8]. Кстати, австралийская парфюмерная промышленность в последнее время широко использует солнцезащитные свойства этих белков при изготовлении губных помад и кремов.
Немецкий ученый Д.Шлихтер и его коллеги полагают, что одна из функций цветной флуоресценции глубоководных видов кораллов - преобразование ультрафиолетового и фиолетово-синего света глубин в более длинноволновый, делающий возможным фотосинтез симбионтов.
В.Вард, один из исследователей, впервые клонировавших GFP из эквореи, высказал предположение, что флуоресценция, в отличие от просто цветной пигментации, - это вариант предупредительной окраски, поскольку она делает кораллы с их стрекательными клетками на щупальцах видимыми для рыб на больших глубинах.
Мы допускаем, что цветные пигменты могут быть ответственны также за фотоактивацию пока не идентифицированных ферментных систем, и в частности за тканевую фоторецепцию. Однако ни одна из этих гипотез не объясняет неодинаковую окраску разных участков тела у многих видов. Например, малиновую (Anemonia sulcata), зеленую флуоресцирующую (A.majano), оранжевую (Zoanthus sp.), синюю (Heteractis crispa) у кончиков щупалец или бирюзовую возле ободка вокруг подошвы (Actinia equina), между тем как все остальное тело - белое, бурое (от симбиотических водорослей зооксантелл), малиновое (A.equina) или с ярко-зеленой флуоресценцией и т.д.
На подводных снимках шельфа в Красном море мы заметили, что на малой глубине многие сидячие формы (губки, кораллы, асцидии и др.) окрашены под цвет водорослевого фона: в зонах преобладания красных, бурых или диатомовых и зеленых водорослей, животные бывают соответственно красными, желто-бурыми и зелеными. Покровительственная окраска или случайные совпадения? Если такое распределение закономерно, то цвет маскирует животных от зрячих хищников или обеспечивающие его белки необходимы для каких-то фотохимических реакций, меняющихся с глубиной. Механизм же можно объяснить просто: пелагические личинки сидячих животных избирательно оседают на поверхность, заселенную одной из этих групп водорослей, которые сами (или обитающие на них бактерии) выделяют привлекающие их вещества (аттрактанты). Однако на подводных снимках, сделанных вне Красного моря, заметить аналогичное распределение цветных сидячих животных нам не удалось.
Статистически доказано: разделение флуоресцентных окрасок кораллов по крайней мере на красный и зеленый варианты отнюдь не случайность. Его поддерживает естественный отбор. Но для чего? Почему? Здесь еще непочатый край работы. Ведь особого прогресса не заметно на протяжении последних десятилетий.
Головокружительная “научная карьера” светящихся и цветных белков
Светящиеся и флуоресцентные индикаторы. От фундаментальной науки не принято ожидать сиюминутного практического выхода. Но светящиеся и цветные белки привлекли внимание сразу же после их открытия. Мы упоминали, что экворин стали применять в качестве индикатора ионов кальция в клетке. Эти ионы присутствуют в самых разных клетках, а повышением их концентрации в цитоплазме запускается множество клеточных процессов: мышечные и немышечные сокращения, синаптическая передача нервных импульсов, всевозможные виды клеточной секреции, иммунные реакции фагоцитов, клеточные деления и т. д. Так что с помощью подобного индикатора можно следить за всеми перечисленными событиями в клетке. Именно светящиеся белки кишечнополостных животных из класса Hydrozoa - экворин и обелин - положили начало новой эре в биофизике клетки.
В 1985 - 1995 гг. гены экворина и нескольких похожих на него фотопротеинов (обелина и т.д.) выделили, а потом клонировали в кишечной палочке. Позже гены экворина и обелина были внедрены в геномы кукурузы, клеток млекопитающих и т.д. Достаточно было добавить к таким клеточным культурам восстановленный субстрат целентеразин, проходящий благодаря своей гидрофобности сквозь клеточные мембраны, чтобы наблюдать в хроническом опыте за свечением, отражающим изменения концентрации цитоплазматического кальция. Подобные эксперименты продолжаются и поныне. Целентеразин для этой цели специально синтезируют. Биотехнологические фирмы поставляют его заказчикам наряду с плазмидами экворинового гена.
Шельф Красного моря.
Здесь, на малой глубине, кораллы окрашены под цвет фона,
создаваемого водорослями [15].Затем генные инженеры и биотехнологи занялись флуоресцирующими белками, точнее, их генами. В качестве генетического маркера первым начали применять ген GFP из эквореи, соединяя его с генами других белков или вводя его матричную РНК во всевозможные клетки. Стало ясно, что таким способом можно сделать видимыми (во флуоресцентном микроскопе) места и скорость образования любых белков, прослеживать рост клеточных клонов, включая патогенные бактерии и раковые опухоли. Удается также наблюдать за размножением в подопытном организме разных вирусов, в том числе тех, которые содержат не ДНК, а РНК, т.е. ретровирусов, например, иммунодефицита человека. К 2002 г. количество работ с применением GFP в качестве генетического маркера превысило 9000!
В последнее время этот маркер стали применять в комплексе с геном белка нестина, характерного для стволовых клеток мозга. Благодаря этому была опровергнута незыблемая, казалось бы, догма о неспособности нервных клеток восстанавливаться. Оказалось, что их самообновление за счет дифференцирующихся стволовых клеток осуществляется даже у взрослых животных, включая человека. Новые нейронные ассоциации у самца канарейки, к примеру, способствуют обучению новой песне. “Муки творчества” влекут за собой включение вновь дифференцирующихся нейронов в ранее сложившийся мозговой “коллектив”! Во флуоресцентном микроскопе на срезах мозга трансгенных мышей с зеленой нестин/GFP меткой прекрасно видны флуоресцирующие пятна в ряде областей [9]. Результаты работ с применением зеленого флуоресцирующего белка эквореи в качестве маркера сейчас просто необозримы *.
* О них можно прочитать в Интернете (http://www.ncbi.nlm.nih.gov/entrez) на ключевое слово GFPПосле того как был клонирован GFP из A.victoria, возникла необходимость обогатить палитру применяемых цветных маркеров, чтобы в одном опыте метить флуоресцентными метками сразу несколько генов. Попытки добиться этого путем мутагенеза гена GFP, пересаженного в кишечную палочку, имели весьма ограниченный эффект. От исходного дикого типа с максимумом свечения в 508 нм появились слабосветящийся “голубой” мутант с максимумом в 440 нм, а также “красный” мутант с максимумом в 526 нм, где свет все равно зеленый.Многоцветная маркировка. Открыв цветные белки, мы получили возможность метить ими клеточные клоны, вводя в них матричные РНК таких белков [7]. Так, наш коллега А.Г.Зарайский инъецировал мРНК двух цветных белков в эмбрион шпорцевой лягушки на стадии восьми бластомеров: красного (от Ricordia yuma) - в левый спинной зачаток и зеленого (от A.majano) - в правый. В результате левая половина выросшего головастика была красной, а правая - зеленой. Их разделяла желтая полоса посредине тела, где смешивались цветные клетки.
Будущее биотехнологии. Как упоминалось в первой части нашей статьи, белки могут светиться не только от ионов кальция, но и от активных форм кислорода, в частности от супероксида. Французский ученый Ж.-М.Бассо с соавторами установил, что у многощетинковых червей из семейства Polynoidae этим радикалом окисляется фотопротеин полиноидин, и тот начинает светиться [10]. Что представляет собой удивительный полиноидин, пока не известно. Клонировать его ген и изучить аминокислотную последовательность белка до настоящего времени не удалось.
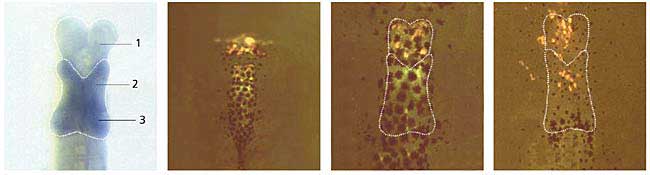
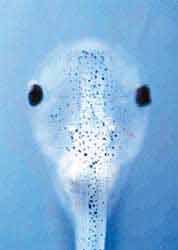
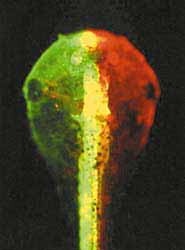
Экспрессия флуоресцентного белка в клетках головного мозга головастика африканской шпорцевой лягушки Xenopus laevis. [16].Фотопротеин из двустворчатого моллюска Pholas dactilys - фолазин - может светиться в присутствии и супероксида, и перекиси водорода, и других активных форм кислорода. В 2000 г. английский биохимик А.К.Кэмпбелл и его коллеги клонировали фолазиновый ген, однако в трансгенных организмах он пока не проявляет себя свечением [11]. У моллюска он образует комплекс с низкомолекулярным субстратом, давно известным, но неидентифицированным; к сожалению, он не похож ни на один из известных люциферинов. Если бы полиноидин и фолазин стали объектами биотехнологии, они нашли бы широчайшее применение в биологических и медицинских исследованиях, ведь содержание и кинетика активных форм кислорода в нервных и других клетках не менее важны, чем цитоплазматическая концентрация ионов кальция. Заметим, что за ее изменением в разных внутриклеточных органоидах можно проследить, используя специальные генетические конструкты. Они должны состоять из генов экворина и того белка, который характерен для той или иной клеточной структуры. Слиянием соответствующих фрагментов ДНК подобного рода конструкты уже изготовлены для многих клеточных структур: митохондрий, их межмембранного пространства, эндоплазматического ретикулума, ядра, синапсов, аппарата Гольджи и т.д.Левый рисунок - структуры головного мозга в проходящем свете (контроль), остальные - при УФ-облучении (эксперимент).
В опытный образец на ранней стадии формирования мозга был введен конструкт, состоящий из гена мутантного флуоресцирующего белка из красной дискосомы и одного из генов, которые контролируют развитие мозговых структур. В результате, по мере экспрессии гена, ответственного за синтез этого белка, появилась зеленая флуоресценция в УФ-свете: сначала - в клетках переднего мозга, позже - промежуточного. Со временем зеленая флуоресценция сменилась на розоватую.
1 - передний мозг, 2 - промежуточный, 3 - средний.
В этом случае изменения концентрации ионов кальция регистрируют при помощи микроскопов с электронным усилением яркости изображения и выводом информации на компьютер. Мы полагаем, что можно добиться одновременной регистрации в разных клеточных структурах - ядре, митохондриях, эндоплазматическом ретикулуме и т.д. Для этого нужны генетические конструкты, которые содержали бы и ген экворина, и гены разноцветных флуоресцентных белков. Тогда цвет свечения экворина будет свой собственный в каждом клеточном органоиде: например в ядре - красным, митохондриях - зеленым, ретикулуме - желтым и т.п. При быстром вращении диска с множеством цветных светофильтров в оптическом пути микроскопа сигналы разного цвета можно направлять для одновременной регистрации на разных каналах электронно-оптической установки. Полагаем, что такие генетические конструкты и установка - дело ближайшего будущего. Эта задача представляет не только теоретический, но и практический интерес. Есть веские основания полагать, что кинетика изменений концентрации Са2+ в каких-то участках нервных клеток характерным образом нарушается при ряде заболеваний центральной нервной системы, например паркинсонизме и болезни Альцгеймера.
Что касается многоцветных меток, то их применение особенно перспективно в так называемом методе индуктивно-резонансного переноса энергии (frequence-resonance-energy transfer - FRET). Впервые освоенный с использованием разноцветных мутантов GFP из эквореи, этот метод позволил наблюдать межбелковые взаимодействия. Чтобы увидеть контакты между белковыми молекулами, один их тип метят сравнительно коротковолновой флуоресцентной меткой, например зеленой (максимум излучения на 508 нм), а другой - длинноволновой, скажем, с красной флуоресценцией (излучением 583 нм), возбуждаемой зеленым светом. Если облучить объект ультрафиолетовым или синим светом, флуоресценция будет зеленой только до тех пор, пока первый из меченых белков не соединится со вторым. Когда эти белки вступят в контакт, флуоресценция станет красной из-за нерадиационной миграции энергии с первой метки на вторую. Так можно изучать взаимодействия антитела с антигеном, вирусного белка и связываемого им белка “хозяина”, молекулярного рецептора с белковым гормоном и т.д. Можно наблюдать разрушение белка ферментами протеазамии *.
* Об этом см. в Интернете (http://www.ncbi.nlm.nih.gov/entrez) на ключевое слово FRET.Другое применение разноцветных белков - автоматическая сортировка клеток по цвету в проточно-капиллярном цитометре. От красного белка все той же красной дискактинии получен мутант, который по мере созревания за 16 ч меняет цвет флуоресценции с зеленого на красный. Если эмбрион пометить геном такого белка, то можно видеть, где ген начал работать раньше (т.е. стал синтезироваться сам белок), а где - позже. Об этом свидетельствует последовательное появление пятен зеленой и красной флуоресценции.
Правда, есть одна методическая трудность. Открытые нами белки, в отличие от зеленого флуоресцирующего белка из эквореи, имеют тенденцию слипаться в уже выделенном виде в димеры, тетрамеры и даже в большие агрегаты. А это затрудняет их применение в методе FRET и может быть даже вредно для исследуемых клеток. Однако в последнее время эта трудность успешно преодолевается - специалисты научились получать неслипающиеся мутантные варианты GFP-подобных белков. Один мономерный белок получен в лаборатории Р.Тсиена путем 33 точковых замен аминокислотных остатков. Другой мутант - “тандемный” - сконструирован в лаборатории С.А.Лукьянова в виде единого гена, слитого из двух одинаковых мономеров. Кодируемый этим димерным геном белок тоже не образует тетрамеров и не слипается в комплексы. Применение таких вариантов весьма перспективно.
Итак, изучение биолюминесценции, а затем и флуоресценции началось с той самой светящейся медузы, которую Форскал назвал эквореей. В те годы, когда экворин впервые начали применять в качестве индикатора ионов кальция, о генной инженерии еще не было и речи. Фотопротеины просто выделяли из свежесобранных медуз или полипов и хранили в морозилке холодильника. Пределом мечтаний ученых было увидеть, как в экспериментах с мышечным волокном внутриклеточная концентрация Са2+ резко возрастает при сокращении или как подобные же ее изменения запускают межнейронную химическую передачу возбуждения в синапсах. Теперь генные инженеры и биотехнологи манипулируют генами флуоресцирующих и цветных белков, чтобы в буквальном смысле наблюдать за ходом внутриклеточных событий. Не исключено, что в будущем им удастся окрашивать разные организмы во все цвета радуги. И это будет передаваться по наследству. * * *
Работа поддержана Российским фондом фундаментальных исследований. Проект 02-04-49717.
Литература
1. Shimomura O., Jonson F.H., Saiga Y. // J.Сell. Сompar. Рhysiol. 1962. V.59. P.223 - 229.
2. Prasher D.C., Eckenrode V.C., Ward W.W. et al. // Gene. 1992. V.111. P.229 - 233.
3. Лабас Ю.А. О механизмах активируемой кальцием биолюминесценции гребневиков // Биофизика живой клетки. 1973. Вып.4. С.83 - 116.
4. Labas Y.A., Gurskaya N.G., Yanushevich Y.G. et al. // Proc. Natl. Acad. Sci. USA. 2002. V.99. P.4256 - 4261.
5. Dove S.G., Hoegh-Guldberg O., Ranganathan S. // Coral reefs. 2001. V.19. P.197 - 204.
6. Wiedenmann J., Elke C., Spindler K.D., Funke W. // Proc. Natl. Acad. Sci. USA. 2000. V.97. №26. P.14091 - 14096.
7. Matz M.V., Fradkov A.F., LABAS Y.A. et al. // Nature Biotechnol. 1999. V.17. №10. P.969 - 973.
8. Salih A., Larkum, A., Cox G. et al. // Nature. 2000. V.408. P.850 - 853.
9. Yamaguchi M, Saito H, Suzuki M, Mori K. // Neuroreport. 2000. V.11. P.1991 - 1996.
10. Bassot J.M., Nicolas M.T. // Histochem. Cell. Biol. 1995. V.104. №3. P.199 - 210.
11. Dunstan S.L., Sala-Newby G.B., Fajardo A.B. et al. // J. Biol. Chem. 2000. V.275. №13. P.9403 - 9409.
12. Shimomura O. The discovery of green fluorescent protein / M.Chalfie., S.Kain. Wiley-Liss., 1998. P.3 - 15.
13. Tsien R. // Ann. Rev. Biochem. 1998. V.67. P.509 - 544.
14. Matz M.V., Lukjanov K.A., Lukjanov S.A. // BioEsseays. 2002. V.24. P.953 - 959.
15. Фридман Д., Мальмквист Т. Чудеса Красного моря. Герцлия (Израиль).
16. Terskich A., Fradkov A., Ermakova G. et al. // Science. 2000. V.290. №24. P.1585 - 1588.