№4, 2005 г.
© Муронец В.И., Плетень А.П.Нобелевская премия получена -
загадки остались
(История открытия инфекционных прионов)В.И.Муронец, А.П.Плетень
Владимир Израилевич Муронец, д.б.н., проф., зав. отд. биохимии животной клетки
НИИ физико-химической биологии им.А.Н.Белозерского МГУ.
Анатолий Петрович Плетень, к.б.н., докторант того же отдела.Губчатая энцефалопатия крупного рогатого скота, или "коровье бешенство", - наиболее известное заболевание последнего десятилетия. Количество публикаций на эту тему в средствах массовой информации можно сравнить с упоминанием проблем, связанных со СПИДом. Хотя совершенно очевидно, что масштабы распространения болезней абсолютно несопоставимы: ВИЧ-инфекцией заражены несколько миллионов людей, а достоверных случаев смерти людей от губчатой энцефалопатии - десятки.
Существуют две основные причины огромного интереса к такому, казалось бы, мало распространенному заболеванию. Одна из них - экономическая. Выявление одного случая коровьего бешенства приводит к уничтожению всего стада крупного рогатого скота и к запрету потребления мясных продуктов из данного региона или целой страны. Такого рода проблемы приводят даже к международным конфликтам и к серьезным экономическим кризисам. Однако важнее вторая причина - загадочность болезни: ее инфекционный агент - белок, названный прионом, его первичная структура абсолютно идентична структуре неинфекционного белка, присутствующего в клетках здорового организма. Различие между белками только в их конформации - укладке аминокислотной последовательности в пространстве.
Понять и доказать, что инфекция переносится белком, было очень сложно, так как в основе всех вирусных заболеваний лежит репликация вирусных нуклеиновых кислот и воспроизводство вируса, а белки такой способностью не обладают. За открытие прионов - нового биологического принципа инфекций - в 1997 г. профессор Калифорнийского университета в Сан-Франциско Стенли Прузинер (Stanley B.Prusiner) получил Нобелевскую премию, однако болезнь осталась столь же загадочной [1-4]. Недаром один из его научных оппонентов так отозвался на награждение Прузинера: "Нобелевская премия - большая честь, но подлинное удовлетворение ученым приносит установление истины".
Что известно о прионовых заболеваниях?
Существует достаточно много болезней, возникающих в результате агрегации внутриклеточных белков, которая приводит к изменению их локализации, а также формы клеток (рис.1). Хорошо известны амилоидозы, когда в различных органах и тканях образуются агрегаты амилоидных белков (амилоидного протеина, легких цепей иммуноглобулинов, транстиретина, b-2-микроглобулина и др.). Сходный процесс протекает в хрусталике глаза при катаракте - слипание кристаллинов. Вероятно, индуцировать агрегацию могут различные воздействия, и прежде всего окислительный и тепловой стрессы. В этом отношении перечисленные заболевания сходны с губчатой энцефалопатией, принципиальным отличием которой служит ее инфекционный характер (трансмиссионность). Таким образом, полное правильное название этой загадочной болезни - трансмиссивная спонгиоформная (похожая на губку) энцефалопатия крупного рогатого скота.
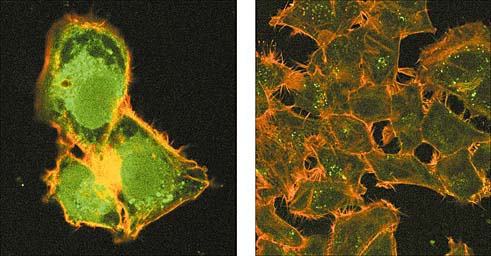
Как передается и развивается губчатая энцефалопатия? В конце 90-х годов прошлого века в Англии зарегистрировали случаи необычной формы нейродегенеративной болезни Крейтцфельда-Якоба. Как правило, это чрезвычайно редкое (1 на 1 млн человек в год) заболевание прионной природы характерно для пожилых людей. Но к середине 1999 г. выявили около 40 больных молодого и среднего возраста. В мозге умерших обнаружили изменения, характерные для губчатой энцефалопатии крупного рогатого скота, и доказали идентичность прионов, выделенных из мозга умерших людей и животных, пораженных коровьим бешенством.Рис.1. Белковые агрегаты, образующиеся в клетках при окислительном стрессе. Фотографии здоровых клеток человека (слева) и клеток, обработанных перекисью водорода. Клетки окрашивали специальными флуоресцирующими антителами и исследовали с помощью конфокального микроскопа. Фото Л.В.Домниной и Е.И.АрутюновойВозникло предположение о возможном заражении людей прионами, содержащимися в тканях больных животных. К этому времени общие представления о развитии прионных болезней и их передаче внутри одного вида уже сложились. В клетках здоровых животных присутствует нормальный прионовый белок (его обозначили PrPC), первичная структура которого полностью идентична инфекционному приону (PrPSc). Однако эти белки сильно различаются конформацией. Инфекционный прион PrPSc очень стабилен и, в отличие от обычных белков, не расщепляется ферментами (протеиназами), плохо растворяется в детергентах и легко агрегирует. Именно эти свойства и обеспечивают инфекционность PrPSc: высокая стабильность позволяет ему преодолеть все барьеры на пути от пищеварительного тракта до клеток мозга, а возникающие высокомолекулярные агрегаты вызывают губчатую энцефалопатию.
Передача прионовых заболеваний внутри одного вида была доказана в нескольких случаях: при употреблении органов и тканей больных животных ("коровье бешенство"), при ритуальном каннибализме (болезнь куру). Но в настоящее время стало ясно, что межвидовые барьеры не абсолютны, и в ряде случаев инфекция передается между особями, принадлежащими к разным видам, в том числе и от животных к человеку. Как правило, при этом болезнь развивается не столь эффективно. Однако нельзя исключить, что при определенном сочетании свойств нормального и инфекционного приона развитие болезни может измениться, в том числе и ускориться. Все этапы прионовых заболеваний - от попадания в организм инфекционного агента до патологических изменений в мозге - до сих пор недостаточно исследованы в силу необычности агента, вызывающего эту страшную болезнь.
Можно ли вылечить губчатую энцефалопатию?
Сегодня методы борьбы с прионовыми заболеваниями основаны на эмпирических подходах. Так, показано, что антибиотик тетрациклин, яд гремучей змеи или антималярийные агенты (хлорпромазин и хинакрин) замедляют болезнь и даже полностью удаляют патогенные прионы из организма [2, 3]. Интересно, что последняя работа сделана в лаборатории Нобелевского лаурета С.Прузинера, который вынужден идти по пути эмпирического поиска средств борьбы с прионовыми заболеваниями. Пессимистические ощущения вызывают и эксперименты по биоэлектронике, выполненные в другой первоклассной лаборатории Сьюзен Лундквист, где исследуются дрожжевые прионы [4]. Очевидно, что ясных путей лечения прионовых заболеваний пока не найдено.
Мы полагаем, что изучение механизмов прионовых заболеваний и поиск методов их лечения должны основываться на успехах в области исследования белков, прежде всего особенностей их укладки, денатурации, агрегации и различных типов посттрансляционной модификации.
В этом плане весьма перспективно изучение роли шаперонов в развитии амилоидозов и прионовых заболеваний. Шаперонами называют белки (или их комплексы), которые помогают полипептидным цепям принимать нативную (природную) конформацию. Шапероны умеют узнавать и связывать неправильно свернувшиеся белки и тем самым препятствуют их агрегации. Некоторые шапероны могут разворачивать неправильно свернутые белки, проявляя так называемую "разворачивающую активность". Очевидно, что шапероны должны играть важную роль в развитии прионоподобных заболеваний. К сожалению, и эта информация весьма ограничена и противоречива.
Так, шапероны, стабилизирующие промежуточные продукты сворачивания полипептидных цепей, способствуют переходу нормального приона в его патогенную форму. Прузинер высказал гипотезу, что превращение нормального клеточного белка PrPc в его стабильную инфекционную форму PrPSc может инициировать молекулярный шаперон. Действительно, в конце 90-х годов С.К.ДебБюрмен и Ф.Эденхофер обнаружили, что различные шапероны человека (Hsp104 и Hsp 60), а также их близкий аналог, выделенный из бактерий (шаперонин GroEL), взаимодействуют с прионовым белком, ускоряя при этом его превращение в инфекционную форму [5, 6]. Позднее Дж.Стокер и Ф.Хартл показали способность GroEL катализировать агрегацию химически денатурированного мутантного приона [7], который в таком состоянии обладал свойствами инфекционного агента.
В то же время имеются данные, что именно нехватка шаперонов в клетке способствует развитию заболевания. Экспериментально показано, что повышенный синтез шаперонов в клетках предотвращает развитие амилоидозов. Однако в клетках, содержащих патогенную форму приона, образование шаперонов не столь активно, как в клетках с нормальным белком [8]. Очень важно, что потеря способности отвечать на стресс синтезом шаперонов служит также признаком стареющих клеток. Возможно, именно этим обусловлен очень длительный инкубационный период прионовых заболеваний (до нескольких десятилетий). Вероятно, после проникновения инфекционных прионов в клетки мозга болезнь не проявляется до тех пор, пока работает система шаперонов, и только после возрастного ослабления их активности развивается губчатая энцефалопатия.
Как изучать губчатую энцефалопатию?
Изучать механизмы возникновения и развития инфекционной (трансмиссионной) губчатой энцефалопатии очень сложно по двум причинам.
Во-первых, трудно выделить нужное для работы количество инфекционного приона. Методы генной инженерии, позволяющие довольно легко получать любое количество нужного белка, в случае инфекционного приона непригодны. Ведь именно превращение обычного белка в патогенный - главная загадка прионов.
Во-вторых, работа с инфекционными прионами требует соблюдения всех правил предосторожности, специального режима экспериментальных исследований. С прионами работать действительно опасно: они очень устойчивы ко многим воздействиям, а способы лечения и вакцинация практически отсутствуют. Все эти причины заставляют исследователей искать удобные и безопасные системы для моделирования прионовых заболеваний.
Существует довольно много неинфекционных амилоидозов, в основе которых лежит формирование так называемых амилоидных фибрилл, сходных по структуре с агрегатами, возникающими в случае прионовых заболеваний. Только у человека более 20 различных белков могут образовывать амилоидные фибриллы. Так, развитие известной болезни Паркинсона связано с агрегацией белка синуклеина, а для болезни Хантингтона и еще пяти сходных заболеваний характерна агрегация белков с полиглутаминовыми фрагментами. Принципиальное отличие этих заболеваний от прионовых амилоидозов - их неинфекционный характер. Следовательно, механизмы возникновения болезни могут быть разными, однако само образование амилоидных структур и дальнейшие патологические изменения оказываются весьма сходны.
Благодаря работам отечественных ученых - С.Г.Инге-Вечтомова, М.Д.Тер-Аванесяна и др. - прионы обнаружены не только у млекопитающих, но и у дрожжей и грибов [9-12]. В природе формирование амилоидных структур далеко не всегда "вредный" патологический процесс. В частности, дрожжевые прионы помогают клеткам адаптироваться к изменениям окружающей среды, т.е. несут некую положительную функцию, в отличие от патогенных прионов млекопитающих. Амилоиды, прионы млекопитающих и дрожжей образуют сходные структуры, но отличаются инфекционностью. У дрожжевых прионов она проявляется в возможности их передачи с цитоплазмой, но не через внеклеточное пространство. Общность некоторых свойств всех типов прионов позволяет использовать дрожжевые прионы как удобную и неопасную модель для исследования прионовых и амилоидных заболеваний.
Достижения в изучении прионов, амилоидов и связанных с ними заболеваний ясно показывают, что в основе всех этих процессов лежит агрегация белков с измененной конформацией. Недавно мы предложили исследовать такой процесс на простых моделях с помощью обычных белков, обладающих теми или иными свойствами прионов. "Искусственные прионы" получали довольно просто: с помощью точечной мутации (замены одного аминокислотного остатка на другой); в результате возникали абсолютно новые пространственные структуры с некоторыми свойствами прионов. Так были получены мутантные формы обыкновенного гликолитического фермента, которые имели повышенную способность к агрегации и связывались с шаперонами так же, как инфекционные прионы.
Однако такой подход хотя и полезен для понимания особенностей агрегации прионоподобных белков, но не совсем адекватен поставленной задаче: ведь последовательность аминокислотных остатков в инфекционном и нормальном прионах абсолютно идентична. Другой подход к индукции сворачивания белков в неправильную конформацию - химическая модификация сульфгидрильных групп. Такие измененные белки связываются с шаперонами, причем необратимо [13-15].
Последнее обстоятельство заслуживает особого внимания, поскольку именно в этом, возможно, и кроется разгадка тайны прионоподобных заболеваний. Хорошо известно, как работают шапероны: они "узнают" неправильно свернутые белки, прочно связывают их, иногда разворачивают, а затем "отпускают", давая полипептиду шанс свернуться правильно. Такую нативную форму шапероны уже не узнают и, следовательно, цикл завершается - денатурированный или неправильно свернутый белок превращается в нативный. Именно этот процесс тормозит развитие болезни, поскольку усиленное образование шаперонов препятствует развитию прионовой патологии.
Совсем иная ситуация будет с модифицированной полипептидной цепью: после аналогичного цикла, осуществляемого шапероном, вновь получится ненативный белок, который повторно свяжется с шапероном. Значит, в основе развития прионовых и амилоидных заболеваний может лежать необратимое блокирование шаперонов.
Из этих наблюдений следуют два важных вывода.
Во-первых, причиной превращения обычного приона в инфекционный может быть неизвестная пока модификация полипепидной цепи, вызванная, например, окислением, фосфорилированием, гликолизированием и другими природными процессами. Любое изменение может индуцировать возникновение структуры, отличной от нативной, в том числе и инфекционного приона, который будет связывать шапероны и стимулировать тем самым образование амилоидных структур. Вероятно, в этом процессе участвуют и другие измененные белки, способные блокировать шапероны, замедляя развитие прионовых болезней. Однако в ходе старения организма (в целом и отдельных клеток) в клетках накапливаются модифицированные полипептидные цепи, которые связывают шапероны и запускают тем самым формирование амилоидных структур. Мы считаем, что инфекционные прионы, блокируя шапероны, должны изменять метаболизм клетки в целом.
Эти процессы чрезвычайно важны для развития прионовых заболеваний, Однако объяснить образование прионовых агрегатов только на их основе очень сложно. Вероятно, сам инфекционный прион не просто способствует агрегации нормальных прионов, а превращает их в инфекционные. Если инфекционный прион связывается с нормальным, а затем их комплекс распадается, то могут возникнуть два инфекционных белка.
С позиций термодинамики такой процесс объясняется следующим образом. В растворе в отсутствие других белков полипептидная цепь принимает строго определенную пространственную структуру. Но если такой полипептид присоединить к какому-то другому белку (или любой полимерной молекуле), он должен свернуться совершенно иначе. Вероятно, именно в этом состоит функция инфекционного приона: в его присутствии обычная полипептидная цепь образует инфекционную структуру. Более того, сам инфекционный прион может служить своеобразной матрицей, на которой идет такое сворачивание. Имитировать эти процессы можно на обычных белках (нативных и модифицированных), а также "подставляя" нативной полипептидной цепи синтетические полимерные молекулы и заставляя ее принимать неправильные формы.
В настоящее время в нашей лаборатории (в содружестве с итальянскими и французскими учеными) созданы модели искусственных шаперонов на основе синтетических полиэлектролитов (совместно с сотрудниками кафедры высокомолекулярных соединений химического факультета Московского государственного университета им.М.В.Ломоносова и шведскими учеными). Проведенные исследования доказали, что в присутствии синтетических полиэлектролитов действительно изменяются термодинамические параметры белков. Значит, для получения неправильно свернутых белков можно использовать синтетические полимеры.
Таким образом, эксперименты с искусственными белковыми системами позволили установить основные механизмы возникновения прионовых заболеваний и амилоидозов, общие закономерности образования белковых агрегатов, лежащие в основе многих патологий, а также наметить пути профилактики и специфической терапии. Апробированные нами белковые системы весьма просты и абсолютно безопасны, что дает возможность существенно расширить круг лабораторий, вовлеченных в изучение прионов.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований
(проект 02-04-48076 и проект, совместный с Белоруссией, 04-04-81038),
а также грантами ИНТАС 03-51-4813 и НАТОLST.СLG. 979533.Литература
1. Prusiner S.B. // N. Engl. J. Med. 2001. V.344. P.1516-1526.
2. Forloni G., Iussich S., Awan T. et al. // Proc. Natl. Acad. Sci. USA. 2002. V.99. P.10849-10854.
3. Korth C., May B.C., Cohen F.E., Prusiner S.B. // Proc. Natl. Acad. Sci. USA. 2001. V.98. P.9836-9841.
4. Scheibel T., Parthasarathy R., Sawicki G., Lin X.M., Jaeger H., Lindquist S.L. // Proc. Natl. Acad. Sci. USA. 2003. Apr.2.
5. DebBurman S.K., Raymond G.J., Caughey B., Lindquist S. // Proc. Natl. Acad. Sci. USA. 1997. V.94. P.13938-13943.
6. Edenhofer F., Rieger R., Famulok M., Wendler W., Weiss S., Winnacker E.L. // J. Virol. 1996. V.70. P.4724-4728.
7. Stockel J., Hartl F.U. // J. Mol. Biol. 2001. V.313. P.861-872.
8. Tatzelt J., Zuo J., Voellmy R., Scott M., Hartl U., Prusiner S.B., Welch W.J. // Proc. Natl. Acad. Sci. USA. 1995. V.92. P.2944-2948.
9. Инге-Вечтомов С.Г., Андрианова В.М. // Генетика. 1970. №6. С.103-115.
10. Kushnirov V.V., Ter-Avanesyan M.D., Didichenko S.A. et al. // Yeast. 1990. V.6. P.461-472.
11. Chernoff Y.O., Lindquist S.L., Ono B., Inge-Vechtomov S.G., Liebman S.W. // Science. 1995. V.268. P.880-884.
12. Kryndushkin D.S., Smirnov V.N., Ter-Avanesyan M.D., Kushnirov V.V. // J. Biol. Chem. 2002. V.277. P.23702-23708.
13. Roitel O., Ivinova O., Muronetz V., Nagradova N., Branlant G. // Biochemistry. 2002. V.41. P.7556-7564.
14. Polyakova O., Asryants R., Roitel O., Branlant G., Muronetz V. // Eur. J. Biochem. 2001. V.268. Suppl 1. P.179.
15. Polyakova O., Roitel O., Asryants R., Branlant G., Muronetz V. // Protein Science, 2005 Mar 1; [Epub. ahead of print].