№ 5, 2000 г.
© П.А.Стунжас, Ф.В.СапожниковЭти удивительные диатомеи
П.А.Стунжас, Ф.В.Сапожников
Павел Антонович Стунжас, кандидат физико-математических наук, старший научный сотрудник лаборатории биогидрохимии Института океанологии им.П.П.Ширшова РАН.
Филипп Вячеславович Сапожников, ведущий инженер лаборатории экологии прибрежных донных сообществ того же института.Известно, что для жизнедеятельности фотосинтезирующим организмам необходимы свет и биогенные соли (биогены). Однако в океане они, как правило, разнесены по глубине (освещены верхние слои, а биогены концентрируются в нижних), поэтому большинство водорослей вынуждены жить при дефиците или одного, или другого фактора. На суше эта проблема решается просто: корни растений — в земле, листья — над ее поверхностью. А в океане надо либо уметь активно перемещаться между верхними и нижними слоями воды, как это делают некоторые виды жгутиковых перидиней, либо регулировать свою плавучесть с помощью газовых вакуолей, подобно некоторым видам синезеленых водорослей (в настоящее время их относят к цианобактериям), либо дожидаться естественных вертикальных движений воды (заметим, очень медленных), что в той или иной степени делают все одноклеточные.
Есть, правда, в океане многоклеточная водоросль Nereocystis, избравшая, как растения на суше, способ механического соединения поверхности воды и грунта. Внешне она напоминает очень вытянутую морковь: длинный (до 100 м и более), утолщающийся кверху и на две трети голый “ствол” в верхней части покрыт “ветвями” и “листьями”. Держаться вертикально этой гигантской “моркови” помогают расположенные в основании “ветвей” и на “листьях” огромные (до 20 см в диаметре) выросты — пузыри с газом. Для океана это, конечно, экзотичный и бесперспективный способ (на 4 км ствол не протянешь), и мы рассказываем о нем лишь для того, чтобы отметить способность многоклеточных водорослей создавать положительную плавучесть с помощью газовых пузырей. Интересно, а может ли это делать одноклеточный фитопланктон — главный производитель органики в океане?
До недавнего времени считалось, что не может, исключение составляли лишь снабженные газовыми вакуолями клетки синезеленых водорослей. Теперь список таких организмов расширился, и в него по праву могут войти некоторые виды диатомовых водорослей, образующих необычные скопления в океане.
Диатомеи в океане
Помимо синезеленых водорослей в состав одноклеточного фитопланктона в основном входят перидинеи, кокколитофориды и диатомеи. Изменения в окружающей среде могут вызвать бурный рост численности любых из них, тем не менее преобладают в океане и соответственно создают бОльшую часть первичного органического вещества диатомеи.
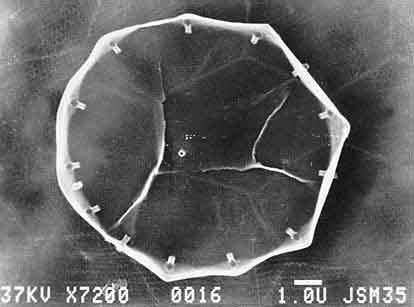
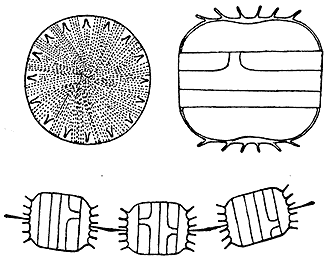
Диатомовые водоросли — сравнительно молодая и резко отличающаяся от прочих группа одноклеточных организмов. Клетки диатомей, размером от менее 10 до более 500 мкм в зависимости от вида, заключены в кремневый панцирь нередко весьма причудливой формы. Панцирь центрической диатомеи напоминает чашку Петри: одна половинка (крышка) чуть больше другой (основания). При делении каждая из них становится крышкой, а размер нового основания уменьшается (но не больше чем на 40% от первоначальной). Пеннатные диатомеи схожего строения, но не круглые сверху, а вытянутые в форме пирожка, вдоль которого проходит шов.
После деления клетки диатомей могут оставаться соединенными между собой в цепочки различной длины. Реже формируются более сложные образования — агрегации из множества цепочек (маты, шарики или комочки, объединенные выделяемой ими же слизью). Если такие агрегации содержат более тысячи клеток, они приобретают макроскопические размеры (более 1мм). У таких макроколоний, как правило, вырабатываются новые, не свойственные отдельным клеткам или простым колониям, приспособительные реакции. Нас заинтересовала, в частности, удивительная плавучесть макроколоний диатомей.
У диатомовых водорослей нет видимых приспособлений для активного передвижения в воде, кремневый панцирь делает их клетки тяжелыми, а для роста им необходим в довольно больших концентрациях кремний — все это уменьшает конкурентоспособность диатомей среди других групп фитопланктона. Возможно, диатомовые сначала были бентосными организмами, в этом случае им ни к чему было всплывать, а уж кремния в иловых водах вдоволь. У живущих в прибрежной зоне современных диатомей нет недостатка ни в освещении, ни в питании, ни в кремнии. А вот как удается обитателям открытого океана не только выживать, но и создавать бо€льшую часть вновь синтезируемого органического вещества? Считалось, что это происходит благодаря тому, что они быстро, опережая другие группы водорослей, размножаются весной, когда в верхнем слое воды после зимнего перемешивания много биогенов, а потом пассивно пережидают неблагоприятные времена либо до будущей весны, либо до случайного заброса биогенов в верхний слой. (В этот период диатомеи сами становятся пищей для зоопланктона и частично бактерий.) После истощения основного запаса биогенов в поверхностном слое начинают преобладать другие виды фитопланктона (перидинеи, кокколитофориды и др.), которые потребляют биогены, образующиеся за счет разложения ранее созданного органического вещества, в том числе и диатомеями.
Однако не все так просто. Диатомеи, как мы знаем, не имеют видимых приспособлений для активного передвижения в воде, но скорость их осаждения зависит от физиологического состояния. Она минимальна в фазе роста и увеличивается при истощении биогенов, включая кремний, хотя из-за его недостатка панцирь становится более тонким. В 1985 г. вышла статья В.Сметачека “О роли погружения в жизненном цикле диатомовых: экологическое, эволюционное и геологическое значение”, где среди прочего описывалось поведение диатомей тропических районов. На апвеллинге у берега или в центре циклонического круговорота на поверхность выходит холодная и сравнительно богатая биогенами вода, и поначалу все идет по описанной схеме: в воде сразу (в тропиках много солнца и поэтому не надо ждать весны) развиваются диатомеи. Вопрос, откуда они там берутся, биологов интересует, как правило, мало — мол, в воде в каком-то минимальном количестве всегда есть характерные для данного района виды. Однако после истощения запаса биогенов (за это время вода прогревается и уносится из области апвеллинга) диатомеи отказываются служить пищей другим членам сообщества и вообще ведут себя необычно.
Происходит это так. Клетки диатомей сцепляются между собой при помощи разного рода выростов или выпускают из себя клейкую слизь, а потом вдруг начинают тонуть. Причем скорость опускания может быть очень большой — до 100 м/сут. Но если случается массовое погружение диатомей, то логично ожидать и такого же массового всплытия. К сожалению, для одиночных клеток и простых колоний диатомей это явление пока не обнаружено, а вот для макроколоний наблюдалось и по крайней мере однажды было очень красиво.
Линия вдоль экватора
В октябре 1994 г. престижный научный журнал “Nature” вышел с фотографией и заголовком статьи “A line in the sea” на обложке [1]. Речь шла о линии в Тихом океане, увиденной и сфотографированной американскими космонавтами с борта станции. Сбылась мечта романтиков, искавших видимую черту, разделяющую Северное и Южное полушария, правда, она оказалась на несколько градусов севернее экватора. К счастью, явление удалось сразу же исследовать с самолета и с научного судна. Эта линия (более 200 км длиной и около 1 км шириной) образовалась во время цветения диатомеи Rhizosolenia, когда наиболее благоприятные условия для ее размножения локализовались вдоль узкого фронта так называемой волны тропической неустойчивости.
Суть явления в том, что довольно часто воды теплого тропического течения, расположенного на несколько градусов севернее экватора, непосредственно соприкасаются с более холодными водами, движущимися в противоположном направлении — вдоль экватора. В результате возникает резкий фронт, когда холодные воды подтекают под теплые и вместе опускаются вниз. Причем скорость опускания холодных вод в данном случае достигала 1см/с, а теплых была гораздо ниже. Таким движением вод сразу же воспользовались диатомеи: на фронте образовалось скопление водорослей очень высокой плотности, при этом концентрация хлорофилла увеличилась до 30 мг/мЗ, т.е. более чем в 100 раз. (Поэтому так хорошо была видна зона цветения с борта космического корабля.) Клубки или маты, в которые соединились диатомеи, не только удерживались в верхних слоях теплой воды, не давая им опускаться, но и увлекаемые холодным течением на сорокаметровую глубину вновь поднимались к поверхности. За время этого вояжа они, видимо, набирались биогенов, так как, оказавшись на свету, начинали интенсивно размножаться.
Как оказалось, миграции Rhizosolenia на глубине 50 м и более — широко распространенное явление в тропических районах океана. В благоприятных условиях нитчатые колонии этой центрической диатомеи образуют хорошо заметные в воде маты размером до 30 см. Выяснилось, что опускающиеся маты содержат меньше нитратов по сравнению с поднимающимися, а поток этих соединений, выносимых водорослями в тропических районах Тихого океана, сравним с подъемом нитратов, который происходит за счет турбулентной диффузии. Так, макроколонии диатомеи Rhizosolenia не только сами решают проблемы поиска биогенов и света в условиях открытого океана, но и обеспечивают другие виды планктона биогенами.
Попробуем оценить скорость миграций водорослей, чего не было сделано в упомянутой работе. Время жизни волны неустойчивости примерно 20 дней, за это время концентрация водорослей увеличилась по сравнению с нижележащим “пустым” слоем в тысячу раз, т.е. каждая клетка должна была претерпеть как минимум 10 делений, что не очень много, так как некоторые виды диатомеи делятся даже 4—6 раз в сутки. Трудно предположить, что каждая клетка может запастись биогенами больше, чем на одно деление. Если считать, что на миграцию у них уходило только половина времени, то скорость должна быть не менее 100 м/сут.
Механизм регулирования плавучести пока не изучен. Авторы статьи предположили, что это — направленный ионный транспорт. На наш взгляд, такой механизм (замена в клетке “тяжелых” ионов “легкими”) неэффективен и, скорее всего, регуляция плавучести осуществляется при помощи газовых пузырьков. Но об этом чуть позже, а пока расскажем о собственных впечатлениях от встреч с необычными колониями диатомовых водорослей.
“Слона-то я и не приметил”
В истории, которая произошла в 1989 г. на судне “Профессор Водяницкий” Института биологии южных морей (Севастополь), участвовал один из авторов (П.А.Стунжас). Рейс был посвящен изучению продуктивности апвеллингов тропической Атлантики. На станции возле мыса Кап-Блан привычный ход работ был нарушен из-за того, что от водной поверхности до глубины 20—30 м появилось множество каких-то бурых скользких чешуек размером 1—2 см. Трудно было отобрать пробу воды в 100 мл, чтобы в ней не попался НПО (неопознанный плавающий объект, названный по аналогии с НЛО), а уж когда опустили сеть для ловли зоопланктона, то ее ячейки тут же забились.
Вскоре мы определили, что НПО — это необычные, очень большие колонии водорослей (как позже выяснилось — диатомеи Thalassiosira parthenia), но к этому времени ведра, полные НПО, были выплеснуты за борт и не осталось ни одного живого образца, только зафиксированная проба. По рассказам очевидцев, в одном из предыдущих рейсов института в этот район они уже встречались с такими объектами и их тоже “не заметили”, осталась только короткая запись в бортовом журнале. Как тут не вспомнить “героя” баснописца в кунсткамере.
Не терпелось исправить допущенную оплошность, однако, вернувшись через два месяца на то же место, мы сразу ничего не нашли. Только спустя сутки произошла долгожданная встреча. На этот раз их было совсем мало, за полчаса удалось наловить всего около ста колоний, но этого было достаточно, чтобы провести исследования.
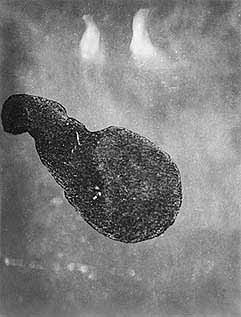
Встреченные в обоих случаях макроколонии Th.parthenia имели вид студенистых буроватых ленточек, которые легко изгибались и скручивались. Самые крупные из них были около 5 см в длину и 2—4 мм в ширину. Под микроскопом можно было разглядеть, что каждая колония состояла из множества (примерно 300 тыс. в колонии длиной 2 см) отдельных клеток размером 8—15 мкм. В отличие от большинства колоний других видов диатомей (в частности, матов Rhizosolenia, состоящих из отдельных цепочек или их клубка) наши подопечные имели более сложную организацию. Они были заключены в хорошо оформленную, плотную оболочку, при повреждении которой (в результате нагревания или старения) из нее выделялось нитевидное содержимое, которое вскоре превращалось в бесформенное облако и пропадало вовсе.
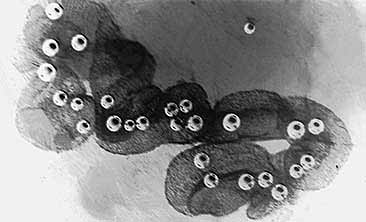
Макроколония диатомеи Thalassiosira parthenia (слева) и ее немногочисленная, но очень красивая спутница, так и оставшаяся неопознанной — НПО.Мы обнаружили, что первичная продукция четырех макроколоний длиной 2 см сопоставима с продукцией, производимой микропланктоном, содержащимся в 1 л окружающей колонии воды (2.4 и 2.6 мг С/м3ч соответственно). Несомненно, макроколонии находились в хорошем физиологическом состоянии. Отметим, что интенсивное развитие на апвеллинге разного рода колоний фитопланктона приводит к значительной неоднородности распределения (сантиметрового масштаба) растворенного кислорода [2]. А как раз поиск такого рода неоднородностей с целью использования их для биологической экспресс-диагностики вод и входил в задачи нашего рейса.Мы предположили, что подниматься к поверхности и удерживаться там макроколониям помогают газовые пузырьки. Наши догадки подтвердились во время дальнейших экспериментов [3]. Плавучесть макроколоний проверялась следующим образом. В сосуд для регистрации акустического рассеяния, которое измерялось узконаправленным эхолотом на частоте 6 МГц, помещали 30—40 колоний. Если сосуд более двух часов находился в темноте, то все колонии опускались на дно. Но примерно через полчаса после включения света, на колониях появлялись микроскопические (40—50 мкм) пузырьки газа, которые приподнимали их, а через час колонии довольно равномерно распределялись по объему сосуда, и в дальнейшем, если на сосуд никак не воздействовали, ничего почти не менялось. При этом пузырьки росли, диаметр самого крупного составлял 0.3 мм. Уже одного этого наблюдения было бы достаточно, чтобы говорить о регулируемой плавучести колоний. В самом деле, пузырек в столбе воды может находиться только в неустойчивом равновесии: при случайном уменьшении глубины гидростатическое давление падает и диаметр пузырька растет, что вызывает увеличение “подъемной” силы и дальнейшее всплывание “пузырька” вплоть до поверхности воды. Если пузырек случайно опустится, то будет продолжать опускаться до дна.
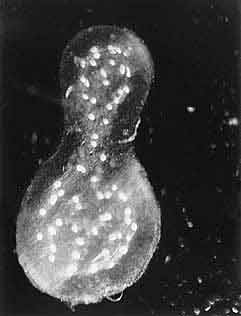
Мы провели дополнительные опыты, которые доказали, что колонии удерживались в столбе воды благодаря регулируемым пузырькам, и оценили некоторые особенности такой регуляции. В сосуде можно было изменять давление воздуха, чем мы и воспользовались. После увеличения давления на 0.2 атм. (что соответствует заглублению на 2м) колонии начали медленно тонуть, но через 10 мин погружение прекратилось! Уменьшение давления на поведении колоний не отразилось, они не всплыли. Этого не могло произойти, если пузырьки были бы пассивными. Подняли давление до 2 атм., и колонии вновь стали погружаться, причем довольно быстро (до 1—2 см/с). Через минуту давление сняли, и две из 40 колоний стали быстро, со скоростью примерно 5 см/с, подниматься, остальные оставались на месте в течение 20 мин, но на них заметно выросли пузырьки. Изменение давления отразилось и на их акустических свойствах. Итак, давление влияло на плавучесть колоний: изменением диаметра пузырьков они, видимо, старались стабилизировать свое положение в столбе воды. Напомним, что пузырьки появлялись на колониях только на свету, а это означает, что наполняющий пузырьки газ — кислород, образующийся при фотосинтезе.
Макроколония Thalassiosira parthenia длиной 4 см в конце шестичасового опыта по определению первичной продукции методом 14С. Хорошо видны пузырьки с кислородом, образовавшимся в процессе фотосинтеза.Заметим, что встреченные в разное время в одном районе океана у Северо-Западной Африки макроколонии Thalassiosira значительно различались между собой по максимальным размерам (от 0.5 до 10 см), по количеству клеток (от 50 до 5000 клеток/мм2), по способу соединения клеток между собой (цепочками или при помощи слизи), по наличию вокруг колонии оболочки (как в нашем случае) или ее отсутствию (во всех других). Кроме того, в лабораторных условиях получить макроколонии этой водоросли никому еще не удавалось. Отсюда можно сделать вывод: или эта водоросль может менять способ объединения клеток в макроколонию, или существует несколько очень близких видов рода Thalassiosira.Им не страшны и полярные льды
До сих пор мы рассказывали о макроколониях диатомей, живущих в тропиках. А что можно сказать о видах, обитающих в высоких широтах Арктики, большая часть которой почти круглый год покрыта дрейфующими льдами? Лед пропускает вниз очень мало света, а его и так не много в высоких широтах, к тому же в Арктике сравнительно низкие концентрации биогенов, включая и кремний. Однако диатомеи приспособились к жизни и в таких условиях.
Свойства и роль льда в жизненном цикле фитопланктона сложны и многообразны [4]. Мы будем говорить только о нижней поверхности льда, где часто концентрация водорослей намного выше, чем в подледном метровом слое воды. Есть здесь и макроколонии диатомей, обладающих положительной плавучестью — это такие студенистые образования, напоминающие по форме и размерам медуз (шар или овал с подобием юбки диаметром до 20 см). Видовой состав “шаров” довольно разнообразен, но доминируют среди них пеннатные Fragilaria. “Шары” видны сквозь молодой гладкий лед. И.А.Мельников насчитывал на 50м2 20—30 колоний.
Если движение льдины медленное, они в основном скапливаются в лунках (полости и каверны) льда. Несомненно, здесь наиболее выгодные для колоний условия: максимальная освещенность и достаточно биогенов. При увеличении скорости движения льдины течение частично вымывает диатомей из лунок и количество их в воде увеличивается. Уже из этих наблюдений следует, что “шары” обладают положительной плавучестью — мало отличающейся от нейтральной, но достаточной, чтобы “шар” не врастал в лед быстро. В Арктике лед нарастает в основном снизу, так как температура воды круглый год отрицательная (от –1.4 до –1.6°С). Способны ли “шары” регулировать свою плавучесть и за счет чего она осуществляется, пока неизвестно, так как изучать эти вопросы in situ очень не просто.
Да здравствуют пузырьки!
Итак, макроколонии обладают плавучестью, несмотря на то, что практически все одиночные клетки фитопланктона, а тем более диатомовые, тяжелее воды. Рассказывая о макроколониях разных видов диатомей, мы объясняли их плавучесть наличием пузырьков с газом, но почти не обосновывали свое утверждение. Сделаем это теперь и проанализируем преимущества этого способа по сравнению с другими, обсуждаемыми в литературе.
Напомним, авторы статьи в “Nature” предположили, что макроколонии регулируют свою плавучесть с помощью направленного ионного обмена. Это утверждение основывается на экспериментах с крупными диатомеями (размер клеток порядка 1 мм), в которых и была обнаружена связь плавучести с ионным составом сока клетки: “тяжелые” ионы Ca, Mg, SO4, NO3 замещались на “легкие” Na, К, Cl, NH4. Однако с физической точки зрения этот довод несостоятелен, так как концентрация “тяжелых” ионов в окружающей среде (морской воде) сравнительно мала, поэтому даже полное их отсутствие при сохранении общей ионной концентрации, влияющей на осмотическое давление, не скажется на плавучести.
Другая попытка объяснить плавучесть макроколоний основывается на известной особенности диатомей накапливать питательные вещества в виде капелек масла, плотность которого меньше воды. Но и этот способ вряд ли используется водорослями для регуляции плавучести. Жиры запасаются как бы на “черный день”, т.е. синтезируются, естественно, в верхних освещенных слоях воды, а потребляются в нижних. Чтобы регулировать плавучесть, все должно быть наоборот.
Об использовании пузырьков с газом диатомеями рода Thalassiosira уже подробно рассказано. Но насколько этот механизм может быть универсальным? На образование газовых пузырьков раньше не обращали внимания, видимо, потому, что количество кислорода, генерируемое одной клеткой, образует пузырек столь малого радиуса, что он будет сразу раздавлен поверхностным натяжением. Другое дело, когда объединяется большое число клеток. Например, в Северном море наблюдались хлопья (агрегаты) морского снега, куда попадало много клеток фитопланктона. На свету при определенных условиях (в частности, высокой концентрации кислорода в воде) в них образовывались микроскопические пузырьки газа, и агрегаты всплывали. На поверхности вОлны сбивали пузырьки, и хлопья начинали тонуть. Но по мере опускания на них снова образовывались пузырьки, и цикл повторялся, и так иногда до 10 дней подряд.
Таким образом, пузырьки могут выполнять роль подводного лифта для макроскопических скоплений не только одноклеточных водорослей рода Thalassiosira. Можно предположить, что они работают и в матах, образованных диатомеей Rhizosolenia. Эффективность такого лифта довольно велика, ведь на, по существу, пустой пузырек действует только выталкивающая сила Архимеда. Даже при малых размерах пузырьков их присутствие значительно увеличивает плавучесть матов. Однако такая регуляция плавучести сопряжена и с некоторыми трудностями: размер пузырьков зависит от глубины (каждые 10 м давление увеличивается на 1 атм.), кроме того ночью и на больших глубинах фотосинтез невозможен. Тем не менее на примере Thalassiosira можно говорить, что пузырьковый транспорт работает по крайней мере до 50—60 м. В Средиземном море эти колонии наблюдались именно на такой глубине и были в хорошем физиологическом состоянии, о чем говорит высокое содержание в них хлорофилла и то, что колонии располагались очень плотным и узким слоем толщиной 2—3 м.
* * *
Биологическая океанология сначала развивалась как описательная наука, но макроколоний как особый объект тогда не заметили. В 60—70-е годы стали собирать материал для количественных сопоставлений. И тут оказалось, что первичная продукция органического вещества примерно в 50 раз меньше его деструкции, чего, конечно, быть не может. Начали искать ошибки в методиках и “хвосты” — неучтенные процессы. Кое-что нашли, в результате оценка деструкции уменьшилась в 2—3 раза, а продукции увеличилась в 5—10 раз. Баланс стал сходиться лучше, но все-таки не до конца. В 90-е годы, наконец, заметили всякого рода скопления диатомей и приступили к изучению их роли в продуцировании органического вещества.
Объективности ради заметим, что на скопления водорослей не обращали внимания по объективным причинам. Описанные макроколонии в больших количествах встречаются, по крайней мере пока, сравнительно редко. Простые скопления клеток часто видны только при непосредственном наблюдении в воде (сейчас их стали изучать при помощи видеокамер), а при сборе сетями скопления разрушаются. При этом оставалось невыясненным главное — способность макроколоний к активным миграциям, что невозможно определить посредством измерения продукции традиционными, т.е. скляночными методами. Будем надеяться, что вскоре баланс сойдется более точно.
Авторы благодарят Д.А.Селивановского за интерес к работе и Российский фонд фундаментальных исследований – за поддержку. Гранты 97-05-65630 и 98-05 —65072.
Литература
1 Yoder J.A., Ackleson S.G., Barber R.A. et al. // Nature. 1994. V.371. № 6499. P. 689—692.
2 Стунжас П.А., Селивановский Д.А. // Океанология. 1999. Т.39. № 3. С. 461—468.
3 Крупаткина Д.К., Селивановский Д.А., Стунжас П.А. // Изв. РАН. Серия биол. 1994. № 3. С. 453—461.
4 Подробнее см.: Мельников И.А. Экосистема арктического морского льда. М., 1989.