№ 5, 2000 г.
№ 5, 2000 г.
© А.Ю.Леин, Л.И.Москалев, Ю.А.Богданов, А.М.СагалевичГидротермальные системы океана и жизнь
А.Ю.Леин, Л.И.Москалев, Ю.А.Богданов, А.М.Сагалевич
Институт океанологии им. П.П. Ширшова РАН, Москва
Прошло более ста лет со времени открытия процесса бактериального хемосинтеза [1] и почти четверть века со дня первого визуального наблюдения горячих гидротермальных проявлений с уникальными биологическими сообществами на дне океана [2]. Выяснилось, что рифтовые экосистемы целиком зависят от геотермальной, а не солнечной энергии [3].
Эти два открытия, разделенные во времени ровно на 90 лет, свидетельствовали об огромной геохимической роли хемоавтолитотрофных (и метанотрофных) микроорганизмов, способных синтезировать органическое вещество, используя углекислоту (и/или метан) в качестве единственного источника углерода, а восстановленные минеральные соединения (H2, H2S, S, S2O32–, SO2, NH3, Fe2+, Mn2+ и др.) — в качестве источника энергии. В 1984 г. внимание исследователей океана привлекли сходные с рифтовыми биологические сообщества, развивающиеся у выходов метановых (и сероводородных) газовыделений и флюидов на активных и пассивных континентальных окраинах океана и даже во внутренних морях, например в Черном. Поля подобных образований получили название “сипы” (от англ. seep — просачиваться). Позднее было предложено фауну, обитающую вокруг выходов низкотемпературных гидротермальных источников на активных и пассивных континентальных окраинах, называть фауной холодных высачиваний, или сиповой, в отличие от гидротермальной рифтовой фауны, обитающей возле высокотемпературных источников.
В основе пищевых цепей рифтовых и сиповых животных лежит главным образом органическое вещество, синтезированное в процессе бактериального хемосинтеза и метанотрофии. При этом микроорганизмы могут быть свободно живущими, прикрепленными (в виде бактериальных обрастаний и матов), а также эндо- и экзосимбионтами.
Практически одновременно российскими и западными исследователями был открыт третий после фото- и хемосинтеза метанотрофный синтез органического вещества за счет использования углерода метана [4]. Это было неожиданно для большинства геохимиков, морских геологов и биоокеанологов.
Международным сообществом ученых при поддержке правительств ведущих стран мира были начаты беспрецедентные по своим масштабам исследования гидротермальных систем океана. С 1977 по 1999 г. в океане обнаружено чуть менее 100 активных и множество реликтовых гидротермальных полей.
Сейчас наступает пора осмысления накопленных фактов и их типизации. Предлагаемая читателям “Природы” статья — одна из таких попыток.
Начнем с того, что рассмотрим гидротермальные системы открытого океана и континентальных окраин в составе общей гидротермальной системы Мирового океана.
Открытый океан. Гидротермальная циркуляция морской воды через изверженные породы океанической коры в “горячих точках”, рифтовых зонах срединно-океанических хребтов и в задуговых спрединговых центрах приводит к физической, химической и биологической эволюции коры и океанов. В этом процессе преобладает адвективный перенос [5] масс вещества, растворов и тепла между глубинными слоями земной коры (а в некоторых случаях даже верхней мантии) и гидросферой.
В предлагаемой классификации в рифтовых зонах срединно-океанических хребтов выделяются две циркуляционные системы — над внутрикоровыми магматическими камерами и глубинная. Последняя открыта лишь после исследований на глубоководных аппаратах “Мир” в Атлантике, на гидротермальном поле Логачев (1995 и 1998 гг.). Флюиды глубинной циркуляционной системы отличаются от обычных относительно высоким содержанием СН4, его гомологов и нефтяных углеводородов.
Континентальные окраины. Глобальный процесс разгрузки термальных вод происходит также на активных и пассивных континентальных окраинах океана. В одних случаях флюиды активных окраин формируются в задуговых спрединговых центрах и в мелководных вулканах, где этот процесс связан с различными изверженными породами или с вулканокластическими отложениями. Состав такого флюида несколько отличается от состава гидротермальных растворов открытого океана, но и в тех и других преобладают соединения серы (H2S, S2O32– и др.).
Вторая группа флюидов активных окраин океана рождается в осадочных толщах зон субдукции и на внутренних склонах глубоководных желобов под действием различных геологических факторов. В составе флюидов из аккреционных призм зон субдукции присутствует значительно больше СН4, чем во флюидах других гидротермальных систем активных континентальных окраин.
Преимущественно метановые флюиды формируются также в гидротермальных системах пассивных континентальных окраин, связанных с грязевым вулканизмом и расположенных в районах лавинной седиментации в океане. Флюиды в этом случае образуются при смешении морской воды и метаморфизованных вод погребенных осадков, содержащих восстановленные соединения, и в первую очередь СН4 и различные углеводороды, которые, как правило, возникают в результате высокотемпературного синтеза из захороненного в осадках органического вещества планктонного происхождения. Заканчивая описание нашей классификации, важно подчеркнуть еще раз, что в открытом океане и в задуговых спрединговых центрах флюиды обогащены H2S, H2 и металлами. Температура этих растворов превышает 360°С.
На дне континентальных окраин, в гидротермальных системах, связанных с осадочными отложениями, температура флюидов на выходе колеблется в пределах 0.01—50°С, редко достигая 100°С. В составе этих растворов преобладает СН4 и его гомологи, присутствуют H2S, H2, NH4. Содержание металлов незначительно.
Рассмотренная классификация гидротермальных систем и закономерное распределение связанных с ними главных компонентов газового состава флюидов важны для оценки роли “первичной” и “вторичной” бактериальной продукции в океане.
Процессы хемосинтеза и метанотрофии с образованием первичной бактериальной продукции происходят у выходов флюидов в “горячих точках”, рифтовых зонах и в задуговых спрединговых центрах за счет энергии окисления восстановленных эндогенных соединений. В процессах вторичного бактериального синтеза органического вещества участвуют восстановленные соединения, поступающие из осадочных пород и возникающие в процессе преобразования захороненного в этих породах органического вещества планктонного генезиса. В течение 1986—1999 гг. (с перерывами) определялись величины первичной бактериальной продукции на активных гидротермальных полях рифтовых зон Тихого океана (гора Осевая, хребет Хуан-де-Фука); во впадине Гуаймас (Калифорнийский залив), бассейнах Манус и Лау (задуговой спрединговый центр Юго-Западной Пацифики) и на четырех полях в рифтовых зонах Атлантического океана (районы 14°45’с.ш., 26°с.ш., 29°с.ш., 36°с.ш.).
Пробы для исследования отбирали с помощью подводных обитаемых аппаратов “Пайсис-4 и -11”, глубоководных обитаемых аппаратов “Мир-1 и -2” и комплекса “Rozett”.
На различных индивидуальных полях величина первичной бактериальной продукции варьировала [6] от 20 до 300 (мг С)(м–2)(сут–1), составляя в среднем около 150 (мг С)(м–2)(сут–1).
Величина вторичной бактериальной продукции реально измерена в районе метанового сипа грязевого вулкана Хаакон Мосби в Норвежском море и составила более 10 (мг С)(м–2)(сут–1). В среднем примерно 23% от количества окисленного СН4 приходится на вторичную бактериальную продукцию. С учетом площади активного сипа было рассчитано7, что вне поверхности дна кальдеры грязевого вулкана окисляется более 100 тыс. м3 СН4 в год.
Бактериальная продукция на дне океана представляет собой дополнительное органическое вещество, количество которого сопоставимо или даже превышает количество планктонного органического вещества, достигшего дна. Добавим, что весь этот материал “свежий”, не прошедший деструкционных преобразований в водной толще.
Рассуждения о малых площадях, занимаемых гидротермальными полями, и о “несущественном” вкладе первичной бактериальной продукции в общий баланс органического вещества в океане верны только в общих чертах. Не следует забывать, что протяженность срединно-океанических хребтов составляет 60—65 тыс. км, а их площадь соизмерима с общей площадью всех материков. Непрерывно осуществляемая, по крайней мере последние 200 млн лет, разгрузка эндогенных флюидов на такой территории и развитие вокруг этих источников биологических сообществ дают весьма существенный вклад в накопление автохтонного органического вещества в открытом океане. Другая зона развития гидротерм — континентальные окраины — имеет протяженность около 350 тыс. км. Иными словами, области, где сконцентрированы гидротермальные системы, охватывают примерно 1/3 площади дна Мирового океана8.
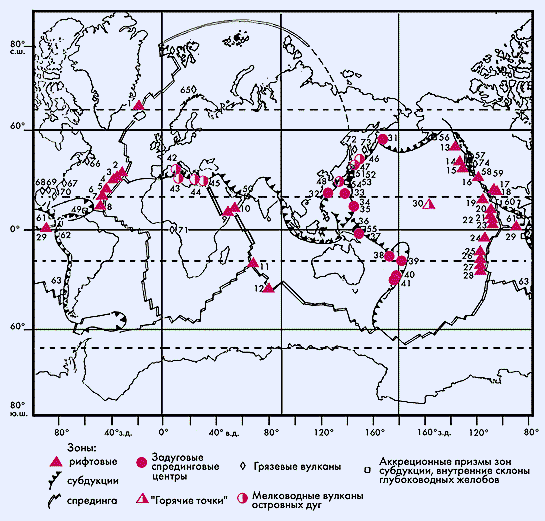
Масштабность процессов бактериального синтеза органического вещества в океане можно проиллюстрировать картой-схемой, на которую нанесены практически все известные на сегодняшний день активные гидротермальные поля в Мировом океане. Читатели “Природы” первыми увидят эту сводку глобального распространения современных оазисов жизни на дне океанов.
Схема распределения рифтовой и сиповой фауны в таком объеме с привязкой к гидротермальным системам океана составлена впервые. Каждому значку на карте соответствует географическое название района. Советские, а затем российские научные экспедиции проводили исследования на 20 активных полях.
Размещение биологических сообществ на активных гидротермальных полях в Мировом океане. Числами обозначены районы гидротермальных проявлений: в открытом океане — рифтовые зоны (1—29), горячие точки (30); на активных континентальных окраинах — задуговые спрединговые центры (31—41), мелководные вулканы островных дуг (42—48), аккреционные призмы зон субдукции и внутренние склоны глубоководных желобов (49—63); на пассивных континентальных окраинах — грязевые вулканы (64—74).1 — хребет Кольбенсей, 2 — Менез-Гвен, 3 — Лаки-Страйк, 4 — Рейнбоу, 5 — Брокен-Спур, 6 — ТАГ (Трансатлантический геотраверс), 7 — Снейк-Пит, 8 — Логачев, 9 — Аденский залив, 10 — рифт Таджура, 11 — тройственная точка Родригес, 12 — плато Амстердам-Св.Павел, 13 —хребет Эксплорер, 14 — хребет Хуан-де-Фука, 15 — хребет Горда, 16 — п-ов Палос Вердес, 17 — бассейн Гуаймас, 18 — Сонорский край бассейна Гуаймас, 19—28 ВТП (Восточно-Тихоокеанское поднятие): 19 — 21°с.ш., 20 — 13°с.ш., 21 — 11°с.ш., 22 — 9°—10°с.ш., 23 — 4°с.ш., 24 — 7°ю.ш., 25 — 17°—19°ю.ш., 26 — 21°—22°ю.ш., 27 — 23°ю.ш., 28 — 26°ю.ш., 29 — Галапагосский спрединговый центр, 30 — вулкан Лоихи,
31 — вулкан Пийпа, 32 — трог Окинава, 33 — трог Огасавара, 34 — Марианский трог, 35 — банка Эсмеральда, 36 — бассейн Манус, 37 — о.Новая Британия, залив Бленч, бухта Матупи, 38 — бассейн Северное Фиджи, 39 — бассейн Лау, 40 — кальдера Брадерс-Арк, 41 — юг дуги Кермадек,
42 — мыс Палинуро, 43 — мыс Мессина и о.Вулькано, 44 — о.Милос, бухта Палеохори, 45 — Восточное средиземноморье, вершина дома Наполи, 46 — о.Янкича, бухта Кратерная, 47 — о.Кунашир, Горячий Пляж, 48 — о.Кюсю, залив Кагосима,
49 — аккреционная призма о.Барбодос, 50 — аккреционная призма Макран, 51 — о.Хонсю, залив Сагами, 52 — Японский и Курильский желоба, 53 — аккреционная призма Нанкай, 54 — каньон Тенриу, восточная часть трога Нанкай, 55 — гора Эдисон, 56 — Алеутский желоб,
57 — Орегонская аккреционная призма, 58 — разлом Сан-Клементе, 59 — залив Монтерей, 60 — мексиканская часть Центрально-Американского желоба, 61 — Коста-Риканская аккреционная призма Центрально-Американского желоба, 62 — Северная часть Перуанского желоба, 63 — центральная часть Чилийского желоба,
64 — хребет Вестнеса, 65 — Хаакон Мосби, 66 — Лаврентийский веер, 67 — континентальный склон Северной Каролины, 68 — каньон Аламинос Мексиканского залива, 69 — континентальный склон Луизианы, Мексиканский залив, 70 — Флоридский эскарп, 71 — Гвинейский залив, 72 — бассейн Дерюгина, 73 — о.Парамушир, 74 — континентальный шельф Северной Калифорнии.
Проводившиеся 23 года исследования гидротермальной фауны позволили установить, что число относящихся к ней видов многоклеточных животных приближается к 500. Подавляющее большинство из них относится к пяти основным типам: членистоногим (35%), моллюскам (34%), кольчатым червям (23%), кишечнополостным (3%) и погонофорам (3%).
Высочайшая концентрация биомассы рифтовых и сиповых сообществ, превышающая 50 кг на 1 м2, не свойственна обычным сообществам животных океанского дна. Почти все виды, обитающие на активных гидротермальных полях (~95%), характерны лишь для зон гидротермалей Мирового океана. Из 207 родов облигатны 47%, из 141 семейства — 18%.
В рифтовых сообществах зарегистрировано вдвое больше видов (475), чем в сиповых (211), хотя доля симбиотрофно питающихся9 приблизительно одинакова (около трети).
Имеющиеся к настоящему времени материалы по гидротермальным системам в корне изменили представление о природе Мирового океана в целом, и в частности об источниках органического вещества в нем. Можно сказать, что со времени открытий, сделанных на научно-исследовательском судне “Челенджер”, океанология не обогащалась за столь короткий срок таким количеством неоспоримых фактов. Полученные в последнее время данные доказывают, что глобальный вынос флюидов из недр Земли активно происходит не только на суше, но и на огромной площади в океане.
Обнаружение многочисленных современных активных, а позднее реликтовых гидротермальных полей на дне океана привело к смене концепции “океан стока” на концепцию “активный океан”. Одна из главных особенностей активного океана — биологическая жизнь на сероводороде и метане. Жизнь, открытая не в лабораторных пробирках, а в масштабе планеты Земля.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Гранты 00-05-64082 и 00-04-48056.
Литература
1 Vinogradsky S.N. // Botanische Zeitung. 1887. №45. S.489—610.
2 Lonsdale P. // Deep—Sea Res. 1997. V.24. P.857—863.
3 Jannasch H.W., Wirsen C.O. // Bio Science. 1979. V.29. P.592—598.
4 Гальченко В.Ф., Галкин С.В., Леин А.Ю. и др. // Океанология. 1988. Т.28. С.1020.
5 Перенос вещества из глубины к поверхности в результате тектонического горизонтального смещения масс океанической коры вдоль срединно-океанических хребтов.
6 Леин А.Ю., Гальченко В.Ф., Пименов Н.В., Иванов М.В. // Геохимия. 1993. №2. С.252—268.
7 Леин А.Ю., Пименов Н.В., Саввичев А.С. и др. // Геохимия. 2000. №3.
8 Fisher A.T. The Quantitative Hydrogeology of Igneous Oceanic Crust: Properties, Driving Forces, and Fluxes // Complex Conference on Multiple Platform Exploration. Vancover, 1999. Р.112.
9 Симбиотрофным принято называть особый тип питания животных за счет жизнедеятельности бактерий внутри или на поверхности тканей животных.