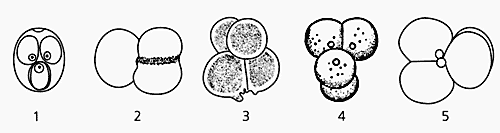№5, 2004 г.
© Сахарова Н.Ю.
Млекопитающие:
эмбрион в личинке
Н.Ю.Сахарова
Наталия Юрьевна Сахарова, кандидат биологических наук, старший научный сотрудник
Института теоретической и экспериментальной биофизики РАН.От Ромула до наших дней
Зарождением новой жизни интересовались еще древние мудрецы, но имели на сей счет самые невероятные представления. Аристотель, например, допускал, что лягушки родятся из ила, а насекомые, черви и прочая мелочь заводятся сами собой в мало-мальски подходящих местах. Алхимики пошли дальше — пытались получить какой-нибудь организм в колбах и ретортах. Сам великий Парацельс дал рецепт изготовления настоящего живого ребенка, правда, очень маленького.
Давно прошли времена алхимиков, поиски гомункулюса сменились естественнонаучными исследованиями. Их объектами стали животные всех крупных и мелких таксонов (изучалось и развитие растительных зародышей), возникла специальная биологическая дисциплина эмбриология. Расцвет ее сравнительно-эволюционной ветви пришелся на последнюю половину XIX—начало XX в. В результате было выявлено сходство в эмбриональном развитии, а значит, и родственные связи, животных, которые, как казалось, исходя из анатомических и физиологических данных о взрослых организмах, эволюционно далеко отстоят друг от друга. Нашлось и место млекопитающим в эволюционном ряду животных, но при этом в их эмбриональном развитии не рассматривался самый ранний период — начальные стадии дробления. Тем не менее можно сказать, что они тоже изучены досконально, им посвящено немало обзоров и монографий, они вошли во многие учебники по биологии развития.
Однако ранний эмбриогенез хранит в себе еще много таинственного и непонятного. Вот высказывания некоторых авторитетных современных исследователей, иллюстрирующие своего рода парадоксы и загадки этого периода развития плацентарных млекопитающих:
– по-видимому, яйцеклетки млекопитающих очень необычны в царстве животных. Ни анимальный ее полюс, ни вегетативный в дробящемся эмбрионе абсолютно не нужны для морфогенеза бластоцисты [1];Эти высказывания заставляют задуматься об интерпретации тех фактов, которые относятся к начальному периоду зародышевого развития. Как же формируется ранний эмбрион млекопитающих?– порядок клеточных делений, приводящих к восьмиклеточной стадии, варьирует у разных эмбрионов, в противоположность тому, что наблюдается в других phyla животных [2];
– до недавнего времени считалось, что ранний (доимплантационный) период важен в основном для образования тканей, обеспечивающих будущий плод питанием и защитой [3];
– термин “эмбрион” в биологии млекопитающих несет оттенок некой двусмысленности: действительно, двуклеточная стадия или бластоциста по сути не являются эмбрионами, эмбрион формируется лишь при гаструляции, а затем развивается в плод [4].
Отличия на фоне общности
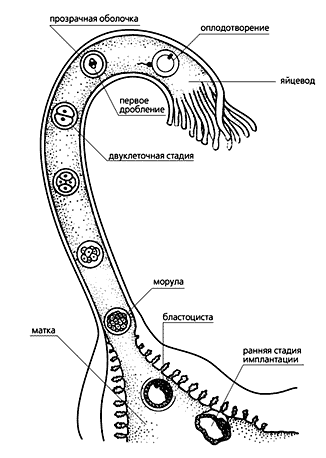
После оплодотворения, которое происходит в то время, когда яйцеклетка на метафазе второго деления созревания находится в верхней части яйцевода, начинается дробление. В этот период зародыш из одноклеточного состояния (зиготы) переходит в многоклеточное путем повторяющихся делений оплодотворенной яйцеклетки. Дробящийся зародыш постепенно перемещается по яйцеводу и в нижней его части достигает восьмиклеточной стадии. Клетки эмбриона, бластомеры, укладываются компактнее (происходит так называемая компактизация), дифференцируются на внутренние и наружные, различающиеся строением и будущей судьбой. Следующие деления приводят к образованию морулы в виде плотного комка клеток, а она превращается в бластоцисту, напоминающую полый пузырек. Ее трофобласт — тонкая стенка пузырька — образован вытянутыми крупными клетками, потомками наружных клеток морулы, а внутри находится небольшое скопление мелких клеток, асимметрично прикрепленных к стенке. Это — внутренняя клеточная масса (ВКМ). На стадии бластоцисты зародыш попадает в матку, выходит из оболочки оплодотворения (zona pellucida) и прикрепляется к стенке матки с помощью трофобласта, который затем станет эмбриональной частью плаценты.
Схема развития зародышей млекопитающих от оплодотворения до имплантации (по Гилберту С., 1993). Дальнейшее развитие посредством делений и дифференцировок внутренних клеток идет по плану, общему для эмбрионов всех позвоночных животных, размножающихся на суше (Amniota), с теми хорошо изученными чертами, которые присущи млекопитающим. Но не об этом пойдет речь, а об особенностях зародыша плацентарных на самых ранних стадиях — в период дробления оплодотворенной яйцеклетки.
У млекопитающих, в отличие от их непосредственных эволюционных предков, яйцеклетки лишены желтка — основного питательного вещества, необходимого для развития зародыша. У других позвоночных в цитоплазме яйцеклетки желток - имеется; полюс, где он сконцентрирован, называют вегетативным, а противоположный — анимальным. Количеством и распределением желтка обусловлены закономерности дробления яйцеклетки у этих животных: бластомеры делятся синхронно, и их положение строго фиксировано.
В безжелтковых (алецитальных) яйцеклетках млекопитающих полярность выражена слабо, а дробление полное и асинхронное. Уже на двуклеточной стадии один бластомер делится быстрее другого, и так продолжается в течение всего периода дробления. Обычно “быстрые” бластомеры поставляют своих потомков в состав внутренней клеточной массы, но бывает, что и в популяцию клеток трофобласта.
У млекопитающих бластомеры на ранних стадиях дробления способны перемещаться. Еще одна примечательная черта — крестообразная фигура, образующаяся после полного завершения второго деления. Возникает она потому, что борозды (границы бластомеров) проходят в двух взаимно перпендикулярных плоскостях.
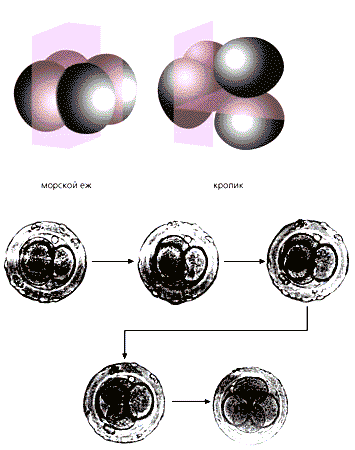
Из-за слабых адгезивных свойств поверхностей бластомеров и перемещений относительно друг друга эти клетки располагаются хаотически, причем в разных эмбрионах по-своему. Это коренным образом отличает восьмиклеточного зародыша млекопитающих от эмбрионов других позвоночных. Асинхронность дробления особенно выявляется у зародышей, в эксперименте лишенных оболочки. На двуклеточной стадии “быстрый” бластомер может делиться в той или иной плоскости, и возникающие в результате клетки располагаются рыхло, образуя клеточные скопления разнообразной формы [1, 2].
Ранний зародыш млекопитающих по характеру развития можно отнести к регуляционным системам, поскольку каждая его клетка поразительно восприимчива к сигналам, идущим от соседей. Это было выявлено в многочисленных экспериментах, выполненных в 60—70-х годах такими классиками эмбриологии, как А.Тарковский, Б.Минтц и др. В раннем зародыше нет четкой определенности в судьбе отдельных бластомеров, их будущее не фиксировано жестко, а зависит прежде всего от взаимного расположения. Потомки любой клетки раннего зародыша способны стать клетками и внутренней клеточной массы, и трофобласта. Более того, бластомеры легко отделяются друг от друга, и каждый из них может дать начало целому организму. Иначе говоря, клетки очень ранних эмбрионов плацентарных идентичны и обладают неограниченными потенциями (т.е. тотипотентны). Обо всем этом говорят результаты опытов, в которых зародыши разделялись на отдельные бластомеры, создавались химеры из клеток разного возраста и в несходных сочетаниях, прослеживалась судьба индивидуальных бластомеров с помощью введения в них метки.
По всем приведенным характеристикам ранние зародыши млекопитающих резко отличаются от таковых других позвоночных. Но ведь одно из доказательств эволюционного родства организмов — сходство в способе дробления, а этого-то и нет в раннем периоде развития зародышей плацентарных животных. Может быть, начальные стадии онтогенеза млекопитающих формировались в эволюции путем возврата к способу, который характерен для далеких предков?Ранние зародыши низших многоклеточных (справа) и млекопитающих. И у первых (метагенетических медуз), и у вторых (мышей, чьи зародыши лишены оболочки оплодотворения) дробление хаотическое (по Захваткину Ю.А., 1949 и Zernicka-Goetz M., 1998).
Еще в конце ХIХ в. И.И.Мечников детально изучил развитие метагенетических гидромедуз — кишечнополостных животных, у которых яйцеклетки не содержат желтка (т.е. алецитальны) и не поляризованы. Оказалось, что дробление у гидромедуз асинхронное и хаотическое, бластомеры способны перемещаться и формировать цепочки, а также отделяться друг от друга, давая начало полноценным особям. Почти все так же, как у млекопитающих!
Если вспомнить, что при дроблении зародышей млекопитающих бластомеры образуют крестообразную фигуру, нужно поискать и ее подобие. Оно найдено: у низших многоклеточных на четырехклеточной стадии дробления образуется “палинтомический” крест, явно напоминающий расположение спор при палинтомическом делении колониальных жгутиковых [5]. Именно по этому общему признаку сделано заключение о несомненной эволюционной связи организмов упомянутых групп.
Таким образом, в своем начальном развитии млекопитающие характеризуются многими особенностями, которые резко отличают их от других позвоночных животных и сближают с низшими многоклеточными, а крестообразное расположение клеток на стадии четырех бластомеров наглядно иллюстрирует сходство с палинтомическим делением у колониальных жгутиковых.
“Палинтомический” крест, образующийся при дроблении у жгутиковых, метагенетических гидромедуз и млекопитающих. У жгутиковых (1, 2) клетки располагаются крестообразно при повторном делении (палинтомии), у гидромедуз (3, 4) и мыши (5) — при втором делении дробления.Следовательно, млекопитающие “используют” способы дробления зародышей низших многоклеточных, лежащих, по современным представлениям, в основании филогенетического древа животных. Тут ко времени напомнить о безжелтковости яйцеклеток млекопитающих — явлении вторичном, произошедшем в процессе эволюции. Известный эмбриолог О.М.Иванова-Казас считает: после исчезновения желтка дробление у них будто бы вернулось на исходные эволюционные позиции и приобрело черты вторичного сходства с дроблением у низших многоклеточных [6]. Например, и у млекопитающих, и у многих гидромедуз на стадии морулы клетки распределяются на внешний и внутренний слои. У гидромедуз в ходе этого процесса, называемого деламинацией, формируются первичные зародышевые листки (экто- и энтодерма), а из них — органы будущего животного. Во время событий, напоминающих деламинацию и происходящих в моруле млекопитающих, клетки тоже дифференцируются на внутренние и внешние [6]. У гидромедуз морула преобразуется в планулу, свободно живущую личинку, у млекопитающих — в бластоцисту. Что же она представляет собой?Личинка и клонирование
Как хорошо известно, отсутствие в яйцеклетке желтка или его малое количество у абсолютного большинства животных связано с наличием в их онтогенезе личиночной стадии. Логично предположить, что ее восстановлением в эмбриогенезе млекопитающих сопровождался возврат их яйцеклеток к алецитальности. Если руководствоваться основным определением эмбриологии как науки, изучающей развитие особи em bryo, т.е. в оболочке, то собственно эмбриональное развитие млекопитающих заканчивается выходом бластоцисты из оболочки оплодотворения (zona рellucida). Сама же бластоциста — это аналог личинки, тело которой составляет внутренняя клеточная масса, а трофобласт служит приспособлением для питания.
С учетом ее дальнейшей судьбы, бластоциста напоминает паразитическую личинку, обладающую специальным приспособлением для развития в теле хозяина — материнском организме.
О том, что млекопитающим свойственно несвободное личиночное развитие, а личиночный период превратился у них в плодный, Г.А.Шмидт говорил более 30 лет назад [7], а С.М.Тилгман совсем недавно высказал мысль о “паразитоподобных” отношениях между эмбрионом и матерью. С помощью трофобласта, который участвует в образовании плаценты, развивающийся организм полностью обеспечивается питанием. Собственно говоря, наиболее важным результатом раннего доимплантацонного развития и считали именно формирование трофобласта как специализированного органа для питания и защиты эмбриона. Это само по себе уже наводит на мысль, что бластоциста никак не может быть эмбрионом ни в каком смысле этого слова. Вероятно, вторичная алецитальность яйцеклетки и должна была привести к появлению стадии личинки в жизненном цикле млекопитающих. Цитируемая уже Иванова-Казас полагает, что установка раннего развития на личинку есть своего рода обходной маневр, после которого развитие возвращается на главную стратегическую дорогу [8]. Что касается млекопитающих, то стадия личинки позволяет продолжить эмбриогенез за счет поступления питательных веществ из тела матери.
Если принять такое объяснение ранних стадий развития млекопитающих, исчезнет оттенок двусмысленности в термине “эмбрион”, о которой упоминает М.Манк [4]. По ее словам, двуклеточная стадия или бластоциста по сути не являются эмбрионами, эмбрион формируется лишь при гаструляции, а затем развивается в плод. В последнее время ранний зародыш принято называть conceptus, т.е. возникший в результате зачатия, а начиная со стадии гаструляции — embryo proper, т.е. собственно эмбрион [9]. Такое различение ведет к представлению о наличии в развитии млекопитающих не последовательных эмбриональных стадий, а двух разных эмбрионов. С этих позиций формирование раннего зародыша следует считать истинным эмбриогенезом, который заканчивается образованием личинки, хорошо известной под именем бластоцисты. Однако неопровержимо и то, что дальнейшее развитие млекопитающих также несет в себе несомненные черты эмбриогенеза, протекающего по образцу, характерному для всех амниот. Этот новый эмбрион (embryo proper) путем прямого развития дает начало плоду, а затем и новорожденному животному.
Как же согласовать наличие стадии личинки и последующего образования нового эмбриона?
Будет крамолой даже подумать, что он развивается из личинки, это категорически противоречит основному постулату эмбриологии, согласно которому невозможно обратить развитие вспять. Личинка никогда не станет вновь эмбрионом, но в ней он может образоваться. Такое явление, хорошо известное как полиэмбриония, детально изучено на примере наездников, перепончатокрылых насекомых, у которых в связи с паразитизмом яйцеклетки стали вторично алецитальными [10]. У наездников морула, возникшая в результате дробления обедненной желтком яйцеклетки, превращается в паразитическую личинку, получающую питательные вещества из организма хозяина через оболочку — трофамнион. Из нескольких соматических клеток тела личинки формируются группы эмбриональных клеток, дающие начало новым личинкам.
Если предположить, что в бластоцисте млекопитающих возникает новый эмбрион, за которым М.Манк только и признавала право называться истинным, то его образование возможно лишь за счет полиэмбрионии. Значит, у этих животных среди клеток, составляющих тело личинки, т.е. среди соматических клеток внутренней клеточной массы бластоцисты, должна быть хотя бы одна группа, способная развиться в новый эмбрион. С таким выводом полностью согласуются результаты экспериментов по исследованию развития химерных зародышей, созданных из генетически различающихся бластомеров [11, 12]. Описание опытов вошло в ставший классическим учебник по биологии развития [13]. На их основании Б.Минтц высказала идею, что начало будущему плоду, а значит, и собственно эмбриону, дает небольшое, четко определенное число клеток из состава внутренней клеточной массы.
Затем гипотеза была проверена в специальных опытах по созданию аллофенных мышей, у которых любой клеточный тип может состоять из фенотипически несходных популяций [11]. Химерные зародыши, образованные из бластомеров, различающихся по генам окраски шерсти, развивались в особи, в чьих шкурках сочетались белый (white) и черный (black) цвета. Проанализировав вероятность сочетаний (W + B)3, Минтц разработала математическую модель происхождения собственно эмбриона из трех клеток внутренней клеточной массы. По этой модели, 75% изначально аллофенных эмбрионов становятся “окончательными мозаиками” и 25% — “немозаиками”, что полностью совпадает с экспериментальными данными. Действительно родились черно-белые мыши, чьи клетки произошли от обоих зародышей — с генами черной и белой окраски. Если бы они развивались только из одной клетки бластоцисты, потомки могли быть либо одного цвета, либо другого; если же из двух клеток, мозаичный рисунок проявился бы только в половине случаев (1WW : 2WB : 1BB). При формировании зародыша из трех клеток вероятность рождения двухцветных животных должна увеличиться до 75% (1WWW : 3WWB : 3WBB : 1BBB ), а из четырех — составить 87.5%. В опытах Минтц 73% химерных зародышей развились в аллофенных мышей, а следовательно, они возникли из трех клеток внутренней клеточной массы.
В других опытах были созданы химерные зародыши из бластомеров, несущих гены трех разных окрасок шерсти. Оказалось, что и в этом случае в 64-клеточной бластоцисте только три клетки служат источником тканей химерных мышей, происходящих от шести родителей. Все три эмбриональных генотипа проявились в пестрой окраске шкурки взрослых мышей [12]. Таким образом, результаты подтверждают гипотезу Минтц, что “только три клетки в ВКМ бластоцисты являются единственными источниками клеток, строящих взрослый организм”.
Итак, согласно Минтц, лишь в трех клетках активируются гены, детерминирующие развитие эмбриона. Именно они формируют три эмбриональных клона, целиком создающие зародыш. Каков механизм выбора этих клеток, пока не известно.
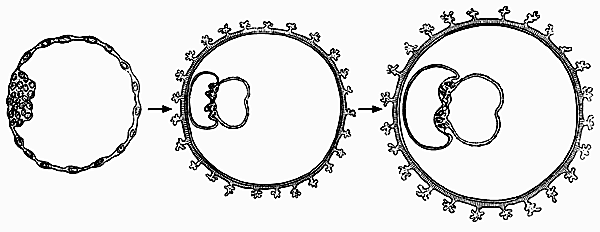
По-видимому, в бластоцисте бывает несколько групп подобных эмбриональных клеток, и тогда рождаются однояйцевые близнецы. В природе такое явление бывает закономерным, свойственным конкретным видам плацентарных млекопитающих, например броненосцам. У вида Tatusia novemcincta постоянно рождаются четыре детеныша одного пола, а у T. hybrida — 12. Это происходит благодаря тому, что все зародыши развиваются из одной бластоцисты, следовательно, в ней у броненосцев существует не меньше четырех клеточных групп, дающих начало эмбрионам.
Но ведь полиэмбриония не что иное, как соматический эмбриогенез — одна из форм бесполого размножения, распространенная в мире низших беспозвоночных. Предположив, что полиэмбриония вовлечена в развитие истинного эмбриона млекопитающих, придется признать, что он формируется внутри бластоцисты путем бесполого размножения. Следовательно, в онтогенезе млекопитающих сочетаются два способа размножения: половой и бесполый. Зачем же нужна такая вставка бесполого размножения? Ведь единственное, что этим достигается — повторение одного и того же организма, т.е. клонирование. Получается, на первый взгляд, невероятное: клонирование включено в нормальное развитие млекопитающих.Возможный способ образования однояйцевых близнецов
из клеток внутренней клеточной массы одной бластоцисты (по Пэттэну, 1959).Идея о наличии личиночной стадии и полиэмбрионии может представить интерес для искусственного клонирования млекопитающих. Если, согласно предлагаемой гипотезе, у них сочетаются половой и бесполый способы размножения, то в геноме яйцеклетки должны быть “записаны” две управляющие программы: первая необходима для контроля за развитием из яйцеклетки личинки, а вторая — из личинки плода. Правда, есть основания думать, что вторая программа не всегда выполняется и онтогенез останавливается на стадии бластоцисты. Вероятно, с этим связан известный медицинский факт: до 40% случаев беременности прерываются еще до имплантации бластоцисты в стенку матки.
Сейчас ясно, что используемые для клонирования донорские ядра, выделенные из соматических клеток, в результате всех манипулирований с ними репрограммируются в цитоплазме безъядерной яйцеклетки-реципиента так, чтобы активировалась первая программа и развилась бластоциста. Вторая же программа срабатывает далеко не во всех случаях. Возможно, в будущем удастся преодолеть эту проблему. Надежду питают данные, полученные при изучении соматического эмбриогенеза у цветковых растений. Под действием специфических факторов роста в культивируемых вегетативных растительных клетках активируется генетическая программа, ведущая их по эмбриональному пути. В результате из нескольких клеток, морфологически ничем не отличимых от остальных, возникает эмбрион, который развивается в новое растение, полную копию того, из которого были взяты исходные клетки.
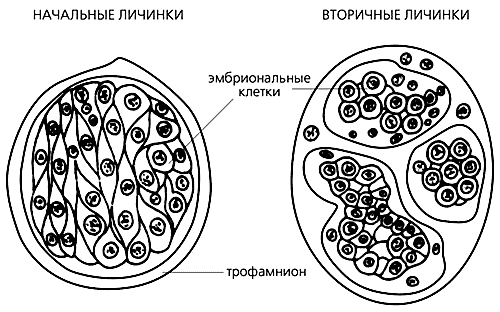
Паразитические личинки перепончатокрылых насекомых — начальные и вторичные.
Последние развиваются из эмбриональных клеток в теле материнской личинки (по Захваткину Ю.А., 1975).Не исключено, что в организме беременной самки тоже вырабатываются специфические факторы роста, которые побуждают соматические клетки к детерминации — выбору пути развития собственно эмбриона. Крайне интересно, что при культивировании эмбриональных стволовых клеток мышей некоторые из них превращаются в оогонии, проходят мейоз, дробление и развиваются до бластоцисты [14]. Это подтверждает возможность активирования в соматических клетках программы образования раннего зародыша. Узнав, какие факторы “запускают” программу соматического эмбриогенеза, мы получим в руки инструмент, с помощью которого можно воздействовать на ядра соматических клеток, пересаженных в пустые яйцеклетки, чтобы пробудить и первую, и вторую программы развития [15].
Геномный отпечаток
Одно из направлений экспериментальной эмбриологии млекопитающих — поиски у них партеногенеза (способности яйцеклетки развиваться без оплодотворения) — напоминает поиски философского камня. Этим в течение столетий занимались средневековые ученые, и все безрезультатно, но зато были сделаны многие открытия, без которых невозможно представить современную науку. Почти все 20-е столетие ученые, поколение за поколением, с увлечением стремились обнаружить возможность развития полноценного организма из яйцеклетки млекопитающих без участия сперматозоида. Ведь бывает же такое в других классах позвоночных! Спонтанно могут развиваться из яйца вполне жизнеспособные особи и у рыб, и у пресмыкающихся, и у птиц, но только не у млекопитающих. Ученые смирились с мыслью, что естественного партеногенеза у этих животных нет, но может быть, его удалось бы вызвать искусственно, как, например, у морских ежей или лягушек. Каким только физическим и химическим воздействиям ни подвергали яйцеклетки млекопитающих, но все напрасно! Итогом долгой и кропотливой работы было заключение: партеногенез в любых его видах у млекопитающих невозможен.
В 1984 г. независимо друг от друга две группы американских ученых пришли к выводу, что материнский и отцовский геномы, передаваемые потомству, качественно дополняют друг друга. Было проведено несколько очень изящных экспериментов по пересадке пронуклеусов (ядер половых клеток с гаплоидным набором хромосом) и искусственному их слиянию, чтобы создать нормальный, двойной, набор хромосом, необходимый для дальнейшего развития яйцеклетки. В результате выяснилось, что и гиногенетические (несущие лишь материнский двойной набор хромосом), и андрогенетические (только с отцовскими хромосомами) эмбрионы не способны полноценно развиваться. Так подтвердилась невозможность партеногенеза у млекопитающих и была доказана обязательность участия в управлении развитием и отцовского, и материнского геномов. Хотя у многих животных оба пронуклеуса эквивалентны, у млекопитающих между ними существуют важные функциональные различия.
Поиски партеногенеза у млекопитающих привели к открытию ядерной памяти, или геномного импринтинга, отпечатка. Именно он обусловливает активность разных генов в отцовских и материнских хромосомах. Иными словами, одни гены работают только тогда, когда наследуются от отца, а другие — если переданы новому организму матерью.
Такие функциональные различия материнского и отцовского геномов имеют особое значение только у плацентарных млекопитающих [16]. Не связано ли это с какими-то характерными чертами их эмбриогенеза и где искать эволюционные корни столь удивительного механизма?
Теперь, исходя из наличия в онтогенезе млекопитающих личиночной стадии, сходной с той, что присуща паразитическим насекомым, нужно попытаться понять, как она возникла и что, в конце концов, представляет собой весь эмбриогенез млекопитающих. Похоже, он состоит из набора определенных “блоков”, которые можно найти в развитии других групп животных, не связанных прямыми эволюционными отношениями. И тогда — это аналогия, вторичное сходство, косвенным доказательством которого может быть геномный импринтинг. Благодаря геномному отпечатку обнаруживается подобие событий, происходящих в развитии определенных групп высших насекомых и высших позвоночных.
Геномный импринтинг был открыт сначала у насекомых (галлиц, ратных комариков и наездников), затем у млекопитающих, а также цветковых растений. В размножении растений, как известно, чередуются половое и бесполое поколения (гаметофит и спорофит). Удивительно, что одним и тем же организмам, независимо от того, к какому миру — животному или растительному — они принадлежат, присущи и геномный импринтинг, и соматический эмбриогенез.
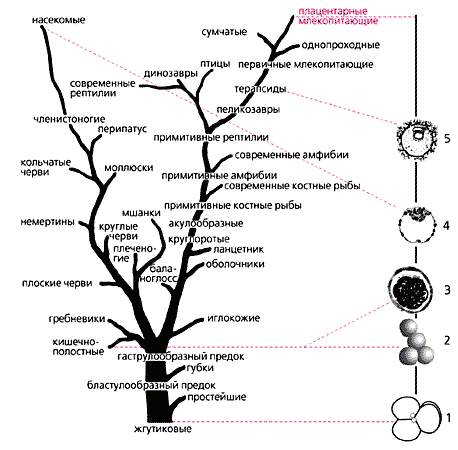
Родословное древо животного мира и схема раннего эмбриогенеза млекопитающих.
1 — стадия четырех бластомеров, расположенных в виде “палинтомического” креста, сходного с расположением спороцитов при палинтомии у колониальных жгутиковых.Существование геномного импринтинга у организмов, принадлежащих не только к разным типам одного царства, но и к разным царствам живого, заставляет вспомнить Э.Жоффруа Сент-Илера, полагавшего, что все разнообразие развития состоит из разных композиций одних и тех же “строительных кирпичиков”.2, 3 — хаотическое дробление и образование морулы, аналогично дроблению у кишечнополостных.
4 — возникновение бластоцисты, паразитической личинки, в которой, как и в паразитических личинках насекомых, идет процесс полиэмбрионии.
5 — формирование собственно зародыша (embryo proper), который будет развиваться аналогично эмбрионам других амниот. (Древо изображено по Вилли К., 1964.)
Млекопитающие, это высшее эволюционное достижение, рассматриваются как продукт последовательной эволюции вторичноротых. Но почему же в раннем онтогенезе столь высоко организованных животных решительно перечеркивается сходство с их прямыми эволюционными предшественниками? Почему он будто возвращается к началу и “попутно” вбирает в себя механизмы, присущие развитию организмов, не связанных с млекопитающими филогенетически? Зачем, будучи представителями высших амниот, млекопитающие “вернулись” к давно “забытому” их непосредственными предками способу развития? Создается впечатление, что и “вставка” в половое размножение бесполого произошла не случайно. Она должна иметь большой смысл для существования млекопитающих как класса. Вероятно, с помощью бесполого размножения в их онтогенез было введено нечто новое, отсутствовавшее у непосредственных предков. Таким новшеством мог быть геномный импринтинг, сопряженный, насколько сейчас известно, с бесполым размножением. А оно, как представляется, входит в качестве определенного периода в онтогенез млекопитающих.
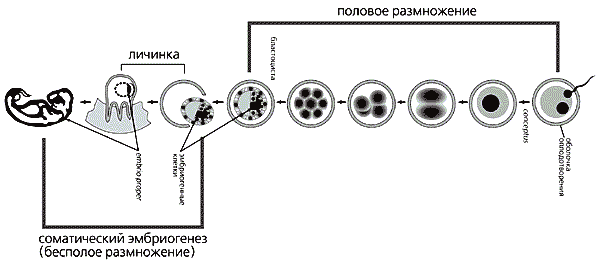
Схема, иллюстрирующая гипотезу о сочетании в онтогенезе млекопитающих
периодов полового и бесполого размножения.С геномным отпечатком связывают активность или пассивность одних и тех же генов в зависимости от передавшего их родителя. Проявляется эта ядерная память по-разному. Например, у листовых комариков и галлиц отцовский набор хромосом полностью разрушается или во время сперматогенеза, или при дроблении. В первом случае отцовские хромосомы не передаются следующему поколению, во втором — вообще отсутствуют в соматических клетках насекомого.
Необычен импринтинг у паразитических перепончатокрылых насекомых — с его помощью регулируется пол потомства. Считалось, что у большинства этих насекомых пол определяется только количеством хромосом: самцы развиваются из гаплоидных, неоплодотворенных яиц, а самки — из диплоидных оплодотворенных. Однако в экспериментах выявилось иное: из диплоидных, но неоплодотворенных яиц, т.е. несущих только материнскую наследственность, всегда развиваются самцы. У одного вида наездников найдена особая хромосома, передаваемая при оплодотворении самцом, которая “убивает” все другие отцовские хромосомы. В результате, даже если от самки получен двойной набор хромосом, в потомстве будут только самцы. Следовательно, эмбрионы развиваются как самки, если несут отцовский набор хромосом, и как самцы, если этот набор потерян. Отцовские хромосомы в следующем поколении становятся материнскими, из чего следует, что в установлении пола у наездников участвует механизм геномного импринтинга [17].
У цветковых растений и млекопитающих импринтингом обусловлены фенотипические отличия материнского и отцовского геномов во время развития, но в гаметах, мужских и женских, они представлены одинаково. Кроме того, есть еще одна параллель. У млекопитающих и цветковых растений в развивающемся зародыше формируются ткани, которые обеспечивают поступление питательных веществ от матери. У млекопитающих это экстраэмбриональные мембраны (в частности, трофобласт), а у цветковых растений — эндосперм. Примечательно. что трофобласт образуется при дроблении самой зиготы, а эндосперм возникает отдельно от нее, при слиянии двух гаплоидных материнских ядер с ядром второго сперматозоида (2m : 1p). Если в избытке отцовские хромосомы (2m : 2p), в эндосперме клеточное деление становится более активным, а если материнские (4m : 1p) деление замедляется.
У мышей экстраэмбриональные ткани гиногенетических и партеногенетических эмбрионов (2m : 0p) тоже развиваются плохо, но сами эмбрионы — почти нормально. Андрогенетические эмбрионы (0m : 2p), наоборот, развиваются слабо, но имеют хорошо развитый трофобласт. В триплоидных человеческих плодах формируется крупная плацента, если дополнительные хромосомы переданы отцом, и слаборазвитая, если они унаследованы от матери.
С геномным импринтингом у млекопитающих связаны и разные патологии. Он имеет большое медицинское значение, так как от него зависит исход многих серьезных генетических заболеваний. Однако нет ясного понимания его функций в нормальном развитии. Существует гипотеза, что геномный импринтинг возник у млекопитающих в связи с конфликтными интересами материнских и отцовских генов в обеспечении питательными веществами, поступающими от матери к потомству [18]. По гипотезе, если тот или иной локус предпочтительно активен в отцовском геноме, питательные вещества будут поступать из материнского организма в увеличенном количестве. Активные материнские гены, наоборот, снизят интенсивность процесса. В целом же предполагают, что оба набора генов соединяются в зиготе так, чтобы дать жизнеспособное потомство, которое будет размножаться, а следовательно, передавать полученные гены следующему поколению. Возможно, мать сохраняет общий контроль над количеством энергии, вкладываемой в отдельных потомков. Отцовские гены запрограммированы на получение как можно большего количества питательных веществ от матери, а материнские — на сдерживание, иначе вся ее энергия растратится только на развивающихся в данную беременность зародышей [18].
Более того, и после рождения, и в течение всей жизни организм постоянно ощущает на себе влияние геномного импринтинга. Яркий пример, иллюстрирующий зависимость активности генов от их родительского происхождения — болезнь, вызываемая потерей одного локуса 15-й хромосомы. Если это материнская хромосома, развивается синдром Ангельмана (некоординированные движения языка при сосании и глотании, частая рвота), а если отцовская, — синдром Прадер-Вилли (слабое сосание, ведущее к значительному нарушению развития).
Известно, что проявление умственных и социальных качеств человека (поведение в социуме) обусловлено разным функционированием определенных генов, передаваемых от отца или от матери своим потомкам в зависимости от пола последних. Судя по исследованиям больных детей с нарушением строения Х-хромосомы, именно в ней находятся гены, от которых зависят познавательные способности человека и его социальное поведение. Так как сын получает только одну, материнскую, Х-хромосому, его умственные и социальные возможности, в первую очередь, наследуются от матери. Иное дело дочери, геном которых формируется Х-хромосомами обоих родителей. Здесь вступает в силу механизм импринтинга: материнские или отцовские копии генов, определяющих интеллект и социальное поведение, выключается.
Есть доказательства, что работающие копии генов, связанных с социальным поведением девочек, находятся на отцовской Х-хромосоме, а материнские копии в дочернем организме молчат. В этом убеждают последствия синдрома Тернера — болезни, при которой у девочек имеется лишь одна Х-хромосома. Если она получена от матери, дочка, в дополнение к другим аномалиям, оказывается и социально неконтактной, ее поведение более характерно для маленького мальчика. Х-хромосома, доставшаяся девочке от отца, обеспечивает поведение, близкое к нормальному. Оказалось, что организм девочки выключает гены на одной из хромосом, а именно на материнской. Следовательно, социальное поведение женщины зависит от отцовской Х-хромосомы, которая всегда наследуется дочерьми. Информация, которая хранится в материнской Х-хромосоме и проявляется у сыновей, не будет считываться в организме дочери [19].
В передаче интеллекта участвуют другие гены на той же хромосоме. Материнские копии некоторых из них активируют рост коры головного мозга, а отцовские, наоборот, тормозят. По-видимому, и девочки, и мальчики получают интеллект от матери [20].
Итак, для млекопитающих геномный импринтинг необычайно важен. Анализ его проявления у других организмов дает возможность предположить, что в процессе эволюции этот механизм был введен в онтогенез млекопитающих вместе с формированием нового эмбриона (embryo proper). А он развивается за счет полиэмбрионии — разновидности бесполого размножения в бластоцисте, аналогичной личинке паразитических насекомых. Природа не тратилась на изобретение, она составила новую композицию из блоков, уже использованных в онтогенезе других организмов. * * *
Литература
1. Zernicka-Goetz M. // Development. 1998. V.125. P.4803—4808.
2. Graham C.F., Deussen Z.A. // J. Embryol. Exp. Morph. 1978. V.48. P.53—72.
3. Pedersen R.A. // Nature. 2001. V.409. ?6819. P.473—475.
4. Биология развития млекопитающих / М.Манк. Пер. с англ. М., 1990. С.10.
5. Захваткин А.А. Сравнительная эмбриология низших беспозвоночных. М., 1949.
6. Иванова-Казас О.М. Эволюционная эмбриология животных. СПб., 1995.
7. Шмидт Г.А. // Арх. анатомии, гистологии, эмбриологии. 1972. Т.63. С.17—18.
8. Иванова-Казас О.М. // Онтогенез. 1997. Т.28. ?1. С.31—40.
9. Johnson M.H., Sellwood L. // Reprod. Feril. Develop. 1996. V.8. P.759—764.
10. Захваткин Ю.А. Эмбриология насекомых. М., 1975.
11. Mintz B. Control Mechanisms in the Expression of Cellular Phenotypes. N.Y., 1970. P.15—43.
12. Markert S.L., Petters R.M. // Science. 1978. V.202. P.56—58.
13. Гилберт С.Ф. Биология развития. М., 1993. Т.1.
14. Hubner K., Fuhrman G., Christenson L.K. et al. // Science Express. 1 May 2003. P.1/10.112.
15. Сахарова Н.Ю., Чайлахян Л.М. // Докл. Акад. наук РФ. 2003. Т.393. ?3. С.426—429.
16. Евсиков А.В. // Онтогенез. 2000. Т.31. ?3. С.178—191.
17. Haig D. // TREE. 1998. V.13. P.380—381.
18. Moore T., Haig D. // TIG. 1991. V.7. ?2. P.45—49.
19. Pagel M. // Nature. 1997. V.387. P.705—708.
20. Keverne B. et al. // Proc. Royal. Soc. L. 1996. V.262. P.689—696.