Июнь 2000 г.
© В.А.РатнерХроника великого открытия: идеи и лица В.А.Ратнер
(Окончание. Начало см.: Природа. 1998. №4, №11)
После того как Дж.Уотсон и Ф.Крик расшифровали молекулярную структуру генов, главным оставался вопрос о генетическом коде. Первым эту проблему сформулировал Г.А.Гамов, что послужило основой для теории генетического кода. Однако попытки его расшифровки оказались почти безрезультатными. Излюбленный прием физиков-теоретиков — рассмотреть множество мыслимых вариантов на все случаи жизни — не принес успеха. Как выразился потом Крик, “к 1959 г. проблема кодирования была в упадке” (Crick F.H.C. // The Genetic code / Cold Spring Harbor Symp. Quant. Biology. 1966. V.31, P.3—9). На вопрос корреспондента газеты, когда будет решена проблема генетического кода, крупнейший советский молекулярный биолог В.А.Энгельгардт ответил: лет через пятьдесят. Задача, действительно, была трудной, но не безнадежной. Вопреки предсказанию, это стало ясно уже через один-два года, когда были нащупаны экспериментальные подходы к ее решению.
“Гены — это атомы наследственности”
Этими словами в 1961 г. американский генетик С.Бензер начал свою итоговую Гарвеевскую лекцию о внутренней структуре гена (Бензер С. Тонкая структура гена // Молекулярная генетика. М., 1963. С.11—32). Проследим, как развивалось это представление.
Г.Мендель: признаки контролируются дискретными наследственными факторами.
В.Иоганнсен: менделевские дискретные факторы “заочно” названы генами.
Т.Морган: гены — неделимые частицы генома, занимающие свое место в хромосоме и на генетической карте, способные контролировать признаки, мутировать и удваиваться.
А.С.Серебровский и Н.П.Дубинин: гены имеют сложную внутреннюю функциональную структуру.
Н.К.Кольцов: гены — это боковые радикалы аминокислот (гипотеза).
Н.В.Тимофеев-Ресовский, К.Циммер, М.Дельбрюк: ген — это гетерогенная макромолекула, имеющая внутреннюю структуру.
Дж.Бидл и Э.Тейтум: гены контролируют структуру белков.
Э.Шредингер: ген — это макромолекула, несущая в себе “шифровальный код”, запись наследуемого свойства.
О.Эвери: материальный носитель генов — ДНК.
Дж.Уотсон и Ф.Крик: ген — это линейная последовательность мономеров двухцепочечной ДНК.
Г.А.Гамов: ген — это линейная последовательность символов четырехбуквенного алфавита нуклеотидов, т.е. генетический текст, кодирующий первичную структуру белка.
Дж.Понтекорво: ген — единица функции (цистрон), мутирования (мутон) и рекомбинирования (рекон).
Продолжил этот восходящий ряд Бензер. Его, еще студента-физика, как и сотни других, впечатлила книга Э.Шредингера “Что такое жизнь с точки зрения физики?”. В 1949 г., поступив в аспирантуру по биологии в Окридж, он получил возможность работать в лабораториях С.Лурии или М.Дельбрюка. По совету Дж.Уотсона, тогда тоже студента, Бензер выбрал лабораторию в Калифорнийском технологическом институте, возглавляемую Дельбрюком, который через год послал своего аспиранта в Париж, в Институт Пастера, к известному специалисту по фагам А.Львову. “Впервые я открыл феномен rII в Париже, но тогда не оценил его значения... Когда я вторично столкнулся с феноменом rII, мой ум был уже подготовлен… Это была система со свойствами высокого генетического разрешения” (Benzer S. Adventures in the rII Region // Phage and the Origins of Molecular Biology / Eds J.Cairns, G.S.Stent, J.D.Watson. Cold Spring Harbor, 1966. P.157—165. ).
Иначе говоря, Бензер обнаружил условную летальность мутаций rII фага T4 при его размножении на различных штаммах бактерии-хозяина. На E.coli B жизнеспособные rII-мутанты ускоренно размножались, убивая бактерий; их можно было размножать и поддерживать. На E.coli K12 мутанты не размножались совсем — были летальны; их можно было скрещивать и отбирать, т.е. вести удобный анализ при помощи классических генетических методов — теста функциональной комплементарности и метода рекомбинационного картирования.
“Разрешающая способность (рекомбинаций — В.Р.) должна была быть достаточной для разделения мутаций, даже если бы они не были локализованы в смежных нуклеотидных сайтах (участках. — В.Р.). Другими словами, это была система, где, как позже выразился Дельбрюк, можно было “привязать генетическую карту к местности”” .
С благословения Дельбрюка Бензер начал строить высокоточную внутреннюю генетическую карту локуса rII фага Т4. За 10 лет (1952—1961) он картировал свыше 1600 rII-мутаций, оценил размер локуса, его физическую длину, получил множество других впечатляющих данных. В последующие годы число исследованных мутаций достигло 2400. За несколько десятилетий локус rII оказался наиболее подробно генетически картированным геном в биологическом мире.
Основные результаты Бензера состояли в следующем:
Таким образом, молекулярное представление о генах приобрело новые очертания. Ген (цистрон) по Бензеру — это непрерывный участок последовательности ДНК (~102—103 пар мономеров), кодирующий белок; состоит из сотен сайтов, мутации которых искажают или уничтожают общую функцию гена, поэтому цистрон как целое — единица функции. Вспомним шутку Н.В.Тимофеева-Ресовского: “Это… не кусочек сливочного масла!” Мутации возможны в любых сайтах вплоть до отдельных нуклеотидов, т.е. ген не единица мутирования (мутон). Мутон равен паре нуклеотидов. Рекомбинации могут разделять смежные пары нуклеотидов, поэтому ген нельзя считать неделимой единицей рекомбинации — реконом, который равен паре нуклеотидов.
- локус rII включает два гена (цистрона) — A и B, которые не перекрываются; в дальнейшем выяснили, что эти гены кодируют белки;
- размер генов ~102—103 нуклеотидных пар (н.п.); потом размер уточнили: rII A ~2400 н.п., белок — 88000 D; rII B ~1200 н.п., белок — 44000 D;
- рекомбинации разделяют участки (сайты) мутирования, между которыми несколько н.п.; впоследствии соседние нуклеотиды удалось разделить;
- локус rII разбит на 308 мутационных сайтов (в дальнейшем — на 370); отдельный сайт в среднем отвечает трем н.п. (по более поздним данным ~10 н.п.). Это достаточно высокая степень заполнения локуса мутациями, почти до уровня кодонов.
Все это прекрасно укладывается в информационно-лингвистический подход. Гены представляют собой не зачатки биологических структур, а напоминают линейные тексты (участки ДНК), записанные при помощи некоторых правил и несущие генетическую информацию о молекулярных структурах и функциях. Они ограничены некоторыми размерами, имеют разные последовательности символов-мономеров. Возможны “бессмысленные” участки, не кодирующие белок или кодирующие испорченные белки. При этом во всех случаях и гены, и некодирующие участки — сегменты молекул ДНК, т.е. построены из того же алфавита четырех нуклеотидов. Поэтому различия между такими текстами не в их физической природе, а исключительно — в последовательностях символов-мономеров. В этом ключ нформационно- лингвистического подхода. Значит, гены — не физические, а информационные единицы наследственности, т.е. предположение Шредингера о “шифровальном коде” действительно подтверждается и материализуется.
Но если гены как последовательности нуклеотидов все же ограниченны по размеру, то они должны быть как-то выделены в геноме, поскольку их основное генетическое качество — дискретность. Далее, должно быть соответствие между линейными порядками нуклеотидов в гене и аминокислот в белке (колинеарное соответствие). Наконец, текст цистрона должен быть тем или иным способом разбит на группы, кодирующие отдельные аминокислоты, кодоны.
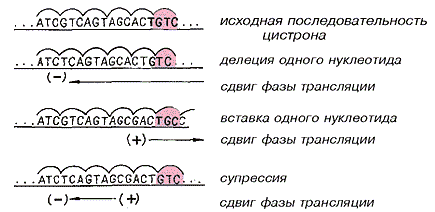
Сдвиг вправо, сдвиг влево — приравнивается к летали
В конце 50-х годов информационно-лингвистический подход в наибольшей мере был присущ Крику. Теоретически он и С.Бреннер поняли, что в цистроне замены символов не нарушают разбиение текста на кодоны, а выпадения (–) и вставки (+) символов сдвигают фазу разбиения соответственно влево или вправо. Комбинирование (–)- и (+)-сдвигов может восстанавливать исходную фазу трансляции. Заметим, что эта идея не оперирует никакими физико-химическими особенностями ДНК, а рассматривает ее абстрактно, как последовательность символов алфавита, разбитую на слова-кодоны.
(+) — вставка символа,
(–) — выпадения символа.
Рассказывают, что для решения этой проблемы Крик с коллегами летом 1961 г. сняли навес для велосипедов в Кембридже, одолжили на время лабораторное оборудование и успешно выполнили свой классический эксперимент, а в конце года опубликовали результаты этой работы в “Nature” (Crick F.H.C., Barnett L., Brenner S., Watts-Tobin R.G. // Nature. 1961. V.192. №4809. P.1227—1232; Крик Ф., Барнетт Л., Бреннер С., Уоттс-Тобин Р. Общая природа генетического кода для белков // Молекулярная генетика. М., 1963. С.33—50.). Они использовали генетическую систему локуса rII фага Т4 Бензера. Несколько ранее в Кембридже Крик и Бензер изучали мутагенез в локусе rII фага Т4, вызванный акридиновыми красителями. Почти все мутации оказались летальными. Хотя данные были не столь очевидны, Крик предположил, что акридины вызывают выпадения и вставки нуклеотидных пар в цепи ДНК. Это и был желанный подход к анализу внутренней организации цистрона. Интуиция не подвела Крика. Его группа получила свыше 120 акридиновых мутаций в небольшом левом сегменте цистрона rII B, вблизи границы с цистроном rII A. Почти все мутации на E. coli K12 были летальными, а 80 из них при комбинировании в одном гене проявили различные свойства восстановления (супрессии) функции гена rII B.
Из этих 80 мутаций можно выделить две группы, (+) и (–), по способности к взаимной супрессии функции гена B при попадании в одну копию гена. При этом:
Информационно-лингвистический подход позволяет очень четко интерпретировать эти данные. Рассмотрим ген (цистрон) как очень длинный текст (предложение), разбитый на короткие слова одинаковой длины (кодоны). Разбиение цистрона на кодоны соответствует фазе считывания текста — трансляции. Рассмотрим влияние вставок и выпадений символов на изменение или восстановление правильной фазы трансляции.
- у двойных мутантов (+ +) и (– –) функция гена B никогда не восстанавливалась;
- у двойных мутантов (+ –) и (– +) во многих случаях исходная функция гена B была восстановлена, но лишь в случаях, когда мутации были на близких расстояниях друг от друга (как потом выяснилось, — не более 50—60 н.п.), а на больших расстояниях такого эффекта не наблюдали;
- у тройных мутантов (– – –) и (+ + +) в нескольких случаях на близких расстояниях трансляция восстанавливалась, но далеко не всегда.
Пусть (+)-мутация связана с внедрением одного или нескольких символов (но не кратно размеру кодона), а (–)-мутация — с выпадением такого же фрагмента. Пусть пары мутаций (+ +)- и (– –) тоже не кратны размеру кодона. Тогда одиночные (+)- и (–)-мутации должны полностью разрушать правильную фазу трансляции “вниз по течению” (т.е. по направлению трансляции). У (+ +)- и (– –)-мутантов правильная фаза трансляции не должна восстанавливаться, а у (+ –)- и (– +)-мутантов — должна.
Если у тройных мутантов, (+ + +) и (– – –), возможно восстановление правильной фазы трансляции, кодоны триплетны или кратны трем (правда, последнее уж очень расточительно!), поскольку происходит сдвиг, кратный трем.
В триплетных кодонах сдвиг влево на один символ с точки зрения фазы трансляции эквивалентен сдвигу вправо на два символа; а сдвиг влево на два символа эквивалентен сдвигу вправо на один. Других вариантов нет. Поэтому должны быть две группы ((+) и (–)) мутантов. В дуплетном коде была бы одна группа, где все мутации восстанавливали бы друг с другом правильную фазу трансляции. В квадруплетном коде было бы более двух групп взаимной супрессии. Поэтому кодоны триплетны без оговорок, а их максимальное число 43=64.
Если в генетическом коде есть некодирующие триплеты (нонсенсы), то при сдвиге образуется как бы новая случайная последовательность кодонов, среди которых могут встретиться и нонсенсы. При этом важен размер участка сдвига: на малых участках нонсенсы могут не встретиться, а на больших встретятся обязательно и станут барьерами трансляции. Если в коде 64 триплетных кодона, а число нонсенсов — два-три, то в этом случае в среднем они должны встретиться на участках сдвига размером больше 20—30 кодонов (40—60 н.п.). Значит, у многих двойных мутантов типа (+ –) и (– +) на участке сдвига фаза трансляции должна восстанавливаться на расстояниях не более 40—60 н.п., когда нонсенсы-барьеры встречаются редко. Это подтверждено экспериментально. Именно поэтому генетический код сильно вырожден и содержит несколько некодирующих триплетов-нонсенсов.
У двойных мутантов (+ –) и (– +) на участке сдвига фаза разбиения на кодоны меняется. Бывшие границы старых кодонов становятся внутренними участками новых, а бывшие внутренние участки — новыми границами. Трансляция при этом не восстанавливается. Это значит, что между кодонами нет никаких специальных разделительных знаков (“запятых”).
Если все кодоны триплетны, то в принципе любой цистрон можно прочитать тремя способами (т.е. в трех фазах). Однако реализуется лишь одна фаза считывания, когда внутри цистрона нет нонсенсов-барьеров. В остальных фазах, как показано выше, нонсенсы встречаются уже на расстояниях порядка 20—30 кодонов. Поэтому, если кодоны не отделены “запятыми” и нет других внутренних способов их выделения (например, “кода без запятых”), то должно существовать фиксированное начало отсчета фазы, т.е. начало трансляции.
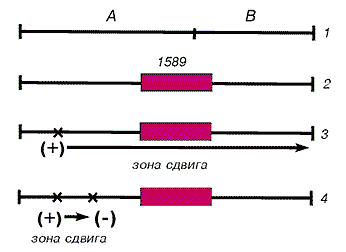
Ранее Бензер нашел тестерную делецию №1589, где выпали правая часть цистрона rII A, левая часть цистрона rII B и все, что было между ними. Крик и коллеги показали, что при этом (+)-мутация в rII А вызывала инактивацию цистрона rII B, а мутация (+ –) в А тоже распространялась в rII B. Следовательно, цистроны А и В сшиты в одной фазе трансляции и считываются слева направо в виде одного общего белкового продукта. Между ними нет препятствий. В нормальном локусе rII цистроны А и В должны быть разделены границей — межцистронным интервалом и двумя знаками: терминальным цистрона А и начальным цистрона В.
В начале 80-х годов, когда описанные зоны цистронов rII A и rII B секвенировали, все эти выводы полностью подтвердились. Даже знак мутаций Крик угадал правильно. Конечно, это чистая случайность, но каков случай! Работа Крика и сотрудников — просто шедевр информационно-лингвистического подхода. К 1994 г. геном фага Т4 прочитали полностью.
при помощи делеции rII 1589.
1 — нормальные цистроны А и В разделены границей трансляции;
2 — делеция 1589 устраняет границу между цистронами, функция В сохранена;
3 — (+)-мутация в А инактивирует функцию В путем общего сдвига вправо фазы трансляции;
4 — добавочная (–)-мутация в А супрессирует (+)-мутацию и восстанавливает функцию цистрона В.
Гонка за лидером
Другая сенсация 1961 г. пришла с неожиданной стороны. Молодой американский биохимик М.Ниренберг и немецкий Х.Маттеи изучали синтез белков в бесклеточной системе трансляции E.coli. Ранее они показали, что для включения меченых аминокислот необходимо участие природной или синтетической РНК, которую потом назвали РНК-посредником, или матричной РНК.
В одном из экспериментов они в качестве матрицы добавили в реакционную смесь гомополимер поли-U. При этом почти в 1000 раз увеличивалось включение меченой аминокислоты фенилаланина в продукт синтеза. Значит кодон UUU кодирует фенилаланин.
Исследователи послали в печать первую работу в марте 1961 г., а затем представили ее на одной из секций V Международного биохимического конгресса, который проходил в Москве 10—16 августа (Ниренберг М.В., Маттеи Г. // Тр. V Международ. биохим. конгресса. Симп.1. 1962. С.211—226). Крик, сразу же поняв, что эта работа сулит “прорыв” в исследовании кода, пригласил Ниренберга повторить доклад на симпозиуме перед большой аудиторией и сопроводил выступление своими положительными комментариями.
Так был дан старт великой “гонке за лидером”. В 1961 г. 34-летний малоизвестный доктор Ниренберг имел небольшую лабораторию в Национальном институте артрита и болезней обмена (г. Бетесда, Мериленд). Начав изучение генетического кода, он сразу же попал в “высококонкурентную среду”. О его работе прослышал крупнейший биохимик, нобелевский лауреат С.Очоа и, поняв, насколько высока ставка, попытался сделать бросок вперед, чтобы опередить Ниренберга. Очоа даже не поехал на конгресс в Москву, а сразу приступил к работе. Силы были неравные: у авторитетнейшего ученого Очоа не было финансовых проблем, кроме того, он владел многими уникальными методами. Но и Ниренберг не собирался сдаваться.
Затем, по словам Крика, “наступила пауза, поскольку было неясно, как продолжать работу и найти порядок оснований в триплетах. Это привело к шквалу теоретических работ, большинство из которых благополучно забыто…”
Я расскажу об одной такой попытке, которая, к моему огорчению, не потрясла мир, но все-таки породила у одного автора определенную уверенность, что такие вещи нужно делать. Этим автором был я сам.
В 1963 г. теоретики располагали:
Ясно, что замены аминокислот должны согласовываться с реальной таблицей кодонов. В принципе, при полноте таких данных можно было бы восстановить относительный порядок оснований во всех кодонах с точностью до одинаковых перестановок символов в таблице, а при неполноте данных — лишь получить относительный порядок оснований только в части кодонов. Чтобы избавиться от неоднозначности за счет перестановок символов, нужны были независимые данные об абсолютном порядке символов хотя бы в одном гетерогенном мутирующем кодоне.
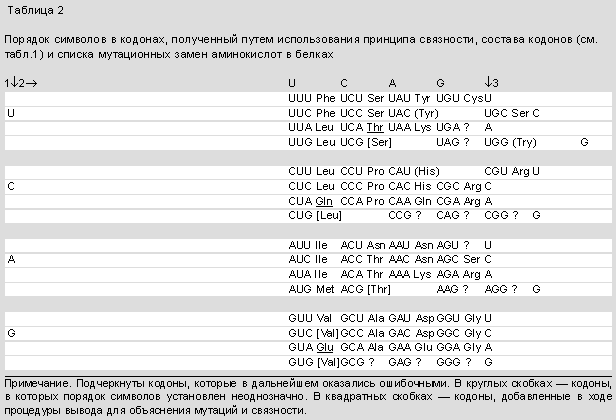
- таблицей состава 48 кодонов; это был явно не полный список, поскольку некоторые кодоны было трудно выявить на фоне других (табл.1);
- списком мутационных замен аминокислот в некоторых природных белках (оболочечном белке вируса табачной мозаики, ВТМ, триптофан-синтетазе E.coli и гемоглобинах человека), также далеко недостаточным. Поскольку некоторые мутации были ненадежны, список часто пересматривали.
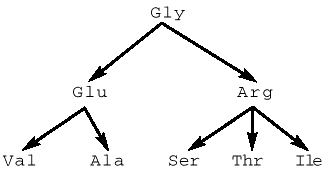
В 1963 г. Ч.Яновский с коллегами опубликовал “мутационную вилку” в 211-м кодоне белка триптофан-синтетазы E.coli:
Некоторые из этих мутантов рекомбинировали между собой:
Arg ґ mVal Ю mGly + Ser.
Яновский показал, что мутант Val имеет маркерную мутацию Ac-(m), локализованную слева от кодона Val за пределами локуса. У мутантов Arg и Val затронуты разные нуклеотиды одного кодона. Сравнение кодонов всех участвующих серий аминокислот между собой и учет перехода левого маркера (Ac-) именно к рекомбинанту Gly приводят к важным следствиям:
Таким образом, Яновский определил порядок двух символов в кодоне Gly.
- все данные совместимы только при предположении, что м-РНК считывается в направлении 5’®3’, если N-конец белка ориентирован влево (этот факт еще не был известен, биохимические данные были неоднозначны);
- мутация Gly®Arg оказалась левее мутации Gly®Glu®Val, т.е. возможные варианты порядка в кодонах Gly могли быть: GGU, GUG, GGA или GAG (помечены участвующие нуклеотиды).
Мутационных данных по-прежнему не хватало, чтобы устранить все неоднозначности. Однако еще раньше, в 1962 г., я заметил, что применительно к структуре генетического кода можно сформулировать одно дополнительное свойство — так называемый “принцип связанности кодовых серий”.
Серией называется группа синонимических кодонов одной аминокислоты. Вспомним, что кодоны опознаются антикодонами т-РНК. Можно предположить, что изоакцепторные фракции т-РНК(переносящие одну аминокислоту) имеют общее эволюционное происхождение и возникают из общей предковой фракции в результате замен нуклеотидов в антикодонах т-РНК. В этом случае все кодовые серии должны быть связанными. Серия называется связной, если, оставаясь в пределах серии, можно из каждого ее кодона получить каждый другой путем последовательных замен отдельных символов. В противном случае серия считается разрывной.
Итак, серии кодонов должны возникать как связные группы. Но может быть, они теряют некоторые кодоны эволюционно и становятся разрывными вторично? Я показал, что это невозможно. Действительно, список кодонов эквивалентен словарю. Если кодоны используются многократно в структурах генов, то утрата любого из них в генетическом коде приводит к множественным дефектам (“дыркам”) в генетических текстах. Такие мутации жестко отсекаются стабилизирующим отбором. Поэтому, каков бы ни был истинный механизм вырождения кода, ни один из когда-либо возникших и часто используемых кодонов словаря не мог быть утерян в ходе эволюции. Значит серии должны быть связны.
Разработанная удобная схема (ее можно назвать алгоритмом) последовательной выбраковки вариантов кода позволила очень быстро сократить их разнообразие до двух-трех. В каждом варианте порядок символов был определен примерно в 20 кодонах. В 1963 г. я послал несколько статей на эти темы, а через год появился большой массив мутационных данных и результаты Яновского о поведении маркера Ac– в рекомбинациях. Мою небольшую статью с последними вариантами кода (где было определено уже до 40 кодонов) и аргументами в пользу направления трансляции (5’®3’) А.Н. Белозерский представил для публикации в “Докладах АН СССР”.
Однако в конце 1964 г. Г.Фунатцу и Х.Френкель-Конрат, отказавшись от предыдущей интерпретации нескольких ключевых мутационных замен аминокислот в оболочечном белке ВТМ, предложили вместо них совершенно другие варианты мутаций (Funatsu G., Fraenkel-Conrat H. // Biochemistry. 1964. V.3. №9. P.1356—1362). Базис моей работы рассыпался, похоронив под обломками все остальные результаты. Я отозвал работу из журнала.
Правда, я тут же переделал работу и получил два новых варианта кодонов. Один из них (А1.1) имел ряд дефектов. Другой (В1.1) был значительно лучше (табл.2). Он очень неплохо совпадал со списком кодонов Ниренберга и его коллег: из 64 триплетов 47 совпали, из-за ошибок в исходных данных и разрывности серии Ser шесть не совпали, состав еще 11 не был определен. Метод себя оправдал. Однако работы в России публикуются медленно — работа вышла только осенью 1966 г. (Ратнер В.А. Генетические управляющие системы. Новосибирск, 1966). Время было упущено. Пиршество победителей уже состоялось.
Что же осталось от этой работы? Во-первых, интерес к проблеме. Во-вторых, понимание, что информационно-лингвистический подход может приносить принципиальные результаты. В-третьих, принцип связности кодовых серий и его эволюционное обоснование. Он выполняется во всех сериях, кроме Ser. В-четвертых, я заметил, что из рекомбинационных данных Яновского вытекает направление трансляции (5’®3’) по матрице. Этот факт еще не был известен и сыграл ключевую роль в выводе вариантов кода.
Пиршество победителей
В 1964 г. М.Ниренберг и Ф.Ледер, преодолев трудности, разработали метод связывания на рибосомах в бесклеточной системе фракций аминоацил-т-РНК с небольшими синтетическими олигорибонуклеотидами заданного порядка оснований. Такие работы, демонстрирующие взаимодействие кодон-антикодон, стали называть экспериментами in vitro. Прежде всего все динуклеотиды оказались неэффективными в связывании т-РНК. Дублетные коды отпали полностью. Метод позволял перепробовать все триплеты по одному.
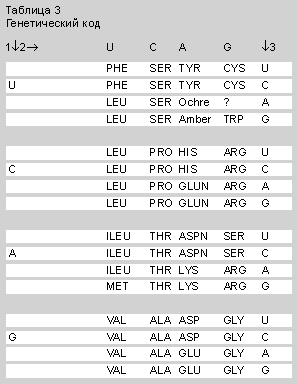
В результате уже через год были испытаны все 64 триплета (Nirenberg M. et al. // The Genetic CODE / Cold Spring Harbor Symp. Quant. Biology. 1966. V.31. P.11—24). Почти все они были признаны кодирующими.
Американский биохимик Г.Хорана пошел другим путем и исследовал матричные свойства синтетических полирибонуклеотидов с повторяющимися парами, тройками, четверками оснований: поли-(UC)n, поли-(AG)n, поли-(UG)n, поли-(UUC)n и др. (Khorana H.G. et al. // Ibid. P.39—49). РНК с повторяющимися парами синтезировали полипептиды с чередующимися аминокислотами, а РНК с повторяющимися тройками кодировали по три раздельных полипептидных гомополимера. Результаты полностью совпали с данными Ниренберга. Известные мутационные замены аминокислот в белках прекрасно соответствовали кодонам. Иначе говоря, данные in vivo полностью совпали с данными in vitro.
2—9 июня 1966 г. в Колд Спринг Харборе, вблизи Нью-Йорка, собрался “съезд победителей” – весь цвет биохимической науки (в основном, американцы). Из СССР был один участник — С.Е.Бреслер из Ленинграда, но без доклада. Таблицу генетического кода, сведенную Криком и представленную как плод коллективного труда, канонизировали как генетический код E.coli.
В своем вступительном докладе Крик сказал:
“Это историческое событие… Оценивая статьи этого симпозиума и оставляя в стороне все сомнительные пункты и оговорки, можно сказать, что открытие генетического кода — это действительно ключ к молекулярной биологии, поскольку он открывает, что два великих языка полимеров — язык полинуклеотидов и язык полипептидов — связаны между собой (выделено мной. — В.Р.). Мы можем быть полностью уверены, что наши общие идеи, такие как гипотеза последовательности (выделено мной. — В.Р.) действительно правильны. После этого для сомневающихся будет очень трудно не принять фундаментальные положения молекулярной биологии, которые мы пытались доказать в течение многих лет”.
Однако в пылу финального успеха фактически были забыты многие выдающиеся теоретики. Гамов не был на симпозиуме; только Крик вспомнил о его роли и в целом отметил роль теоретиков: “Тем не менее теория принесла определенную пользу, будучи время от времени путеводителем для экспериментаторов… Может быть, более важно, что дискуссии о кодировании фокусировали внимание на проблеме и сделали ее более реальной для людей, которые в ином случае просто бы о ней не знали”.
Итак, словарь языка кодонов был определен полностью. Проблема генетического кода нашла свое экспериментальное решение. Структурно-функциональный базис молекулярной биологии получил прочное обоснование. Несмотря на все превратности судьбы, это был и грандиозный успех информационно-лингвистического подхода. Период экспериментальной дешифровки кода успешно закончился. Началось теоретическое осмысление найденных закономерностей. Настало время построения основ теории молекулярно-генетических систем управления, теории генетического языка и др. Но это уже другая история.
В 1968 г. Ниренберг, Хорана и Холли стали лауреатами Нобелевcкой премии по физиологии и медицине за расшифровку генетического кода и его функции в синтезе белка.