№6, 2002 г.
№6, 2002 г.
№6, 2002 г.
№6, 2002 г.
© Г.М.Дымшиц
Сюрпризы митохондриального генома
Г.М. Дымшиц
Григорий Моисеевич Дымшиц, доктор биологических наук, профессор кафедры молекулярной биологии Новосибирского государственного университета, заведующий лабораторией структуры генома Института цитологии и генетики Сибирского отделения РАН. Соавтор и редактор четырех школьных учебников по общей биологии.Со времени обнаружения в митохондриях молекул ДНК прошло четверть века, прежде чем ими заинтересовались не только молекулярные биологи и цитологи, но и генетики, эволюционисты, а также палеонтологи и криминалисты, историки и лингвисты. Такой широкий интерес спровоцировала работа А.Уилсона из Калифорнийского университета. В 1987 г. он опубликовал результаты сравнительного анализа ДНК митохондрий, взятых у 147 представителей разных этносов всех человеческих рас, заселяющих пять континентов. По типу, местоположению и количеству индивидуальных мутаций установили, что все митохондриальные ДНК возникли из одной предковой последовательности нуклеотидов путем дивергенции. В околонаучной прессе вывод этот интерпретировали крайне упрощенно — все человечество произошло от одной женщины, названной митохондриальной Евой (и дочери и сыновья получают митохондрии только от матери), которая жила в Северо-Восточной Африке около 200 тыс. лет назад. Еще через 10 лет удалось расшифровать фрагмент ДНК митохондрий, выделенный из останков неандертальца, и оценить время существования последнего общего предка человека и неандертальца в 500 тыс. лет назад [1].Сегодня митохондриальная генетика человека интенсивно развивается как в популяционном, так и в медицинском аспекте. Установлена связь между рядом тяжелых наследственных заболеваний и дефектами в митохондриальных ДНК. Генетические изменения, ассоциированные со старением организма, наиболее выражены в митохондриях. Что же представляет из себя геном митохондрий, отличающийся у человека и других животных от такового у растений, грибов и простейших и по размеру, и по форме, и по генетической емкости? Как работает и как возник митохондриальный геном у разных таксонов? Об этом и пойдет речь в нашей статье.
У всех эвкариот — будь это малярийный плазмодий, мельчайший одноклеточный паразит, разрушающий эритроциты человека, или сам человек, гигантская свободноживущая клетка амеба протей, микроскопическая колония дрожжей или гриб, имеющий многокилометровый мицелий, эфемерные насекомые поденки или тысячелетние секвойи — у всех генетическая информация содержится не только в хромосомах клеточного ядра, но и в митохондриях — самовоспроизводящихся полуавтономных органеллах клетки, имеющих собственный геном. В то время как ядерный геном представляет собой совокупность линейных молекул ДНК гаплоидного набора хромосом, митохондриальный геном — одну или несколько кольцевых (редко линейных) молекул ДНК (мтДНК). В исключительных случаях эвкариотические клетки не содержат митохондрий, например некоторые паразитирующие в кишечнике анаэробные амебы.
Митохондрии называют энергетическими станциями клетки. Помимо наружной гладкой мембраны они имеют внутреннюю мембрану, образующую многочисленные складки — кристы. В них встроены белковые компоненты дыхательной цепи — ферменты, участвующие в преобразовании энергии химических связей окисляемых питательных веществ в энергию молекул аденозинтрифосфорной кислоты (АТФ). Такой “конвертируемой валютой” клетка оплачивает все свои энергетические потребности. В клетках зеленых растений помимо митохондрий есть еще и другие энергетические станции — хлоропласты. Они работают на “солнечных батареях”, но тоже образуют АТФ из АДФ и фосфата. Как и митохондрии, хлоропласты — автономно размножающиеся органеллы — также имеют две мембраны и содержат ДНК.
В матриксе митохондрий, кроме ДНК, находятся и собственные рибосомы, по многим характеристикам отличающиеся от эвкариотических рибосом, расположенных на мембранах эндоплазматической сети. Однако на рибосомах митохондрий образуется не более 5% от всех белков, входящих в их состав. БOльшая часть белков, составляющих структурные и функциональные компоненты митохондрий, кодируется ядерным геномом, синтезируется на рибосомах эндоплазматической сети и транспортируется по ее каналам к месту сборки. Таким образом, митохондрии — это результат объединенных усилий двух геномов и двух аппаратов транскрипции и трансляции. Некоторые субъединичные ферменты дыхательной цепи митохондрий состоят из разных полипептидов, часть которых кодируется ядерным, а часть — митохондриальным геномом. Например, ключевой фермент окислительного фосфорилирования — цитохром-с-оксидаза у дрожжей состоит из трех субъединиц, кодируемых и синтезируемых в митохондриях, и четырех, кодируемых в ядре клетки и синтезируемых в цитоплазме. Экспрессией большинства генов митохондрий управляют определенные гены ядер.
Размеры и формы митохондриальных геномов
К настоящему времени прочитано более 100 разных геномов митохондрий. Набор и количество их генов в митохондриальных ДНК, для которых полностью определена последовательность нуклеотидов, сильно различаются у разных видов животных, растений, грибов и простейших. Наибольшее количество генов обнаружено в митохондриальном геноме жгутикового простейшего Rectinomonas americana — 97 генов, включая все кодирующие белок гены, найденные в мтДНК других организмов. У большинства высших животных геном митохондрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и два для рРНК (для большой субъединицы рибосом 16S рРНК и для малой 12S рРНК). У растений и простейших, в отличие от животных и большинства грибов, в митохондриальном геноме закодированы и некоторые белки, входящие в состав рибосом этих органелл. Ключевые ферменты матричного полинуклеотидного синтеза, такие как ДНК-полимераза (осуществляющая репликацию митохондриальной ДНК) и РНК-полимераза (транскрибирующая геном митохондрий), зашифрованы в ядре и синтезируются на рибосомах цитоплазмы. Этот факт указывает на относительность автономии митохондрий в сложной иерархии эвкариотической клетки.
Геномы митохондрий разных видов отличаются не только по набору генов, порядку их расположения и экспрессии, но по размеру и форме ДНК. Подавляющее большинство описанных сегодня митохондриальных геномов представляет собой кольцевые суперспирализованные двуцепочечные молекулы ДНК. У некоторых растений наряду с кольцевыми формами имеются и линейные, а у некоторых простейших, например инфузорий, в митохондриях обнаружены только линейные ДНК [2].
Как правило, в каждой митохондрии содержится несколько копий ее генома. Так, в клетках печени человека около 2 тыс. митохондрий, и в каждой из них — по 10 одинаковых геномов. В фибробластах мыши 500 митохондрий, содержащих по два генома, а в клетках дрожжей S.cerevisiae — до 22 митохондрий, имеющих по четыре генома.
Митохондриальный геном растений, как правило, состоит из нескольких молекул разного размера. Одна из них, “основная хромосома”, содержит большую часть генов, а кольцевые формы меньшей длины, находящиеся в динамическом равновесии как между собой, так и с основной хромосомой, образуются в результате внутри- и межмолекулярной рекомбинации благодаря наличию повторенных последовательностей (рис.1).
Рис 1. Схема образования кольцевых молекул ДНК разного размера в митохондриях растений.
Рекомбинация происходит по повторенным участкам (обозначены синим цветом).В митохондриях большинства организмов (кроме высших животных) часть кольцевых молекул ДНК присутствует в виде олигомеров, которые можно разделить на три класса: линейные; кольцевые, имеющие контурную длину, кратную длине мономерных колец; цепные, катенаны, состоящие из топологически связанных, т.е. продетых друг в друга, мономерных колец (рис.2). Так, в единственной митохондрии простейших из отряда кинетопластид, включающего эндопаразита человека — трипаносому, содержатся тысячи кольцевых молекул ДНК. У Trypanosoma brucei имеются два типа молекул: 45 одинаковых макси-колец, каждое из которых состоит из 21 тыс. пар нуклеотидов, и 5.5 тыс. идентичных друг другу мини-колец по 1000 пар нуклеотидов. Все они, соединяясь в катенаны, образуют переплетенную сеть, которая вместе с белками формирует структуру, называемую кинетопластом.
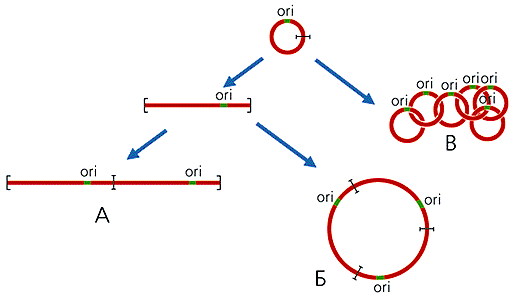
Рис 2. Схема образования линейных (А), кольцевых (Б), цепных (В) олигомеров мтДНК.
ori — район начала репликации ДНК.Размер генома митохондрий разных организмов колеблется от менее 6 тыс. пар нуклеотидов у малярийного плазмодия (в нем, помимо двух генов рРНК, содержится только три гена, кодирующих белки) до сотен тысяч пар нуклеотидов у наземных растений (например, у Arabidopsis thaliana из семейства крестоцветных 366924 пар нуклеотидов). При этом 7—8-кратные различия в размерах мтДНК высших растений обнаруживаются даже в пределах одного семейства. Длина мтДНК позвоночных животных отличается незначительно: у человека — 16569 пар нуклеотидов, у свиньи — 16350, у дельфина — 16330, у шпорцевой лягушки Xenopus laevis — 17533, у карпа — 16400. Эти геномы сходны также и по локализации генов, большинство которых располагаются встык; в ряде случаев они даже перекрываются, обычно на один нуклеотид, так что последний нуклеотид одного гена оказывается первым в следующем. В отличие от позвоночных, у растений, грибов и простейших мтДНК содержат до 80% некодирующих последовательностей. У разных видов порядок генов в геномах митохондрий отличается.
Высокая концентрация активных форм кислорода в митохондриях и слабая система репарации увеличивают частоту мутаций мтДНК по сравнению с ядерной на порядок. Радикалы кислорода служат причиной специфических замен Ц®Т (дезаминирование цитозина) и Г®Т (окислительное повреждение гуанина), вследствие чего, возможно, мтДНК богаты АТ-парами. Кроме того, все мтДНК обладают интересным свойством — они не метилируются, в отличие от ядерных и прокариотических ДНК. Известно, что метилирование (временная химическая модификация нуклеотидной последовательности без нарушения кодирующей функции ДНК) — один из механизмов программируемой инактивации генов [3].
Репликация и транскрипция ДНК митохондрий млекопитающих
У большинства животных комплементарные цепи в мтДНК значительно различаются по удельной плотности, поскольку содержат неодинаковое количество “тяжелых” пуриновых и “легких” пиримидиновых нуклеотидов. Так они и называются — H (heavy — тяжелая) и L (light — легкая) цепь. В начале репликации молекулы мтДНК образуется так называемая D-петля (от англ. displacement loop — петля смещения). Эта структура, видимая в электронный микроскоп, состоит из двуцепочечного и одноцепочечного (отодвинутой части Н-цепи) участков. Двуцепочечный участок формируется частью L-цепи и комплементарным ей вновь синтезированным фрагментом ДНК длиной 450—650 (в зависимости от вида организма) нуклеотидов, имеющим на 5'-конце рибонуклеотидную затравку, которая соответствует точке начала синтеза Н-цепи (ori H). Синтез L-цепи начинается лишь тогда, когда дочерняя Н-цепь доходит до точки ori L. Это обусловлено тем, что область инициации репликации L-цепи доступна для ферментов синтеза ДНК лишь в одноцепочечном состоянии, а следовательно, только в расплетенной двойной спирали при синтезе Н-цепи. Таким образом, дочерние цепи мтДНК синтезируются непрерывно и асинхронно (рис.3).
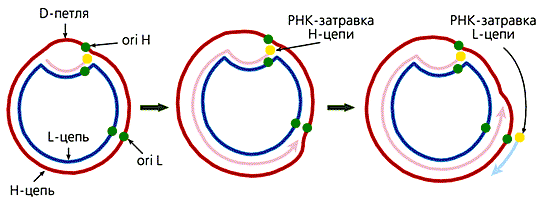
Рис 3. Схема репликации мтДНК млекопитающих.
Сначала формируется D-петля, затем синтезируется дочерняя Н-цепь,
потом начинается синтез дочерней L-цепи.В митохондриях общее число молекул с D-петлей значительно превышает число полностью реплицирующихся молекул. Обусловлено это тем, что у D-петли есть дополнительные функции — прикрепление мтДНК к внутренней мембране и инициация транскрипции, поскольку в этом районе локализованы промоторы транскрипции обеих цепей ДНК.
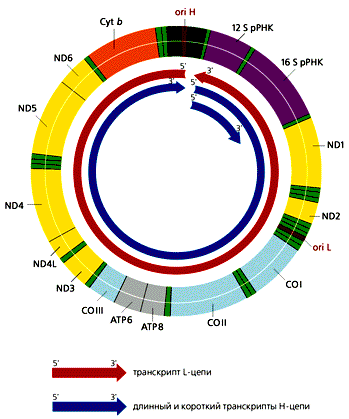
В отличие от большинства эвкариотических генов, которые транскрибируются независимо друг от друга, каждая из цепей мтДНК млекопитающих переписывается с образованием одной молекулы РНК, начинающейся в районе ori H. Помимо этих двух длинных молекул РНК, комплементарных Н- и L-цепям, формируются и более короткие участки Н-цепи, которые начинаются в той же точке и заканчиваются на 3'-конце гена 16S рРНК (рис.4). Таких коротких транскриптов в 10 раз больше, чем длинных. В результате созревания (процессинга) из них образуются 12S рРНК и 16S рРНК, участвующие в формировании митохондриальных рибосом, а также фенилаланиновая и валиновая тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются транслируемые мРНК, к 3'-концам которых присоединяются полиадениловые последовательности. 5'-концы этих мРНК не кэпируются, что необычно для эвкариот. Сплайсинга (сращивания) не происходит, поскольку ни один из митохондриальных генов млекопитающих не содержит интронов.
Рис 4. Транскрипция мтДНК человека, содержащей 37 генов. Все транскрипты начинают синтезироваться в районе ori H. Рибосомные РНК вырезаются из длинного и короткого транскриптов Н-цепи. тРНК и мРНК образуются в результате процессинга из транскриптов обеих цепей ДНК. Гены тРНК обозначены светло-зеленым цветом.Сюрпризы митохондриального генома
ND1—ND6, ND4L — гены субъединиц НAД-H-дегидрогеназного комплекса;
СОI—COIII — гены субъединиц цитохром-с-оксидазы;
ATP6, ATP8 — гены субъединиц AТФ-синтетазы
Cyt b — ген цитохрома b.Несмотря на то, что в геномах митохондрий млекопитающих и дрожжей содержится приблизительно одинаковое количество генов, размеры дрожжевого генома в 4-5 раз больше — около 80 тыс. пар нуклеотидов. Хотя кодирующие последовательности мтДНК дрожжей высоко гомологичны соответствующим последовательностям у человека, дрожжевые мРНК дополнительно имеют 5'-лидерную и 3'-некодирующую области, как и большинство ядерных мРНК. Ряд генов содержит еще и интроны. Так, в гене box, кодирующем цитохромоксидазу b, имеется два интрона. Из первичного РНК-транскрипта автокаталитически (без участия каких-либо белков) вырезается копия большей части первого интрона. Оставшаяся РНК служит матрицей для образования фермента матуразы, участвующей в сплайсинге. Часть ее аминокислотной последовательности закодирована в оставшихся копиях интронов. Матураза вырезает их, разрушая свою собственную мРНК, копии экзонов сшиваются, и образуется мРНК для цитохромоксидазы b (рис.5). Открытие такого феномена заставило пересмотреть представление об интронах, как о “ничего не кодирующих последовательностях”.

Рис 5. Процессинг (созревание) мРНК цитохромоксидазы b в митохондриях дрожжей.
На первом этапе сплайсинга образуется мРНК, по которой синтезируется матураза,
необходимая для второго этапа сплайсинга.При изучении экспрессии митохондриальных генов Trypanosoma brucei обнаружилось удивительное отклонение от одной из основных аксиом молекулярной биологии, гласящей, что последовательность нуклеотидов в мРНК в точности соответствует таковой в кодирующих участках ДНК. Оказалось, мРНК одной из субъединиц цитохром-с-оксидазы редактируется, т.е. после транскрипции изменяется ее первичная структура — вставляется четыре урацила. В результате образуется новая мРНК, служащая матрицей для синтеза дополнительной субъединицы фермента, последовательность аминокислот в которой не имеет ничего общего с последовательностью, кодируемой нередактированной мРНК (см. таблицу).
Происходит это за счет сдвига рамки считывания на число нуклеотидов, не кратное размеру триплета (в данном случае на четыре). Новая белковая субъединица, необходимая для работы фермента, образуется в митохондриях паразита только тогда, когда он попадает в организм холоднокровной мухи и нуждается в окислительном фосфорилировании для получения большого количества молекул АТФ. Если трипаносома живет в организме теплокровных млекопитающих, ей достаточно АТФ, образующейся в процессе гликолиза.
Впервые обнаруженное в митохондриях трипаносомы редактирование РНК широко распространено в хлоропластах и митохондриях высших растений. Найдено оно и в соматических клетках млекопитающих, например, в кишечном эпителии человека редактируется мРНК гена аполипопротеина.
Наибольший сюрприз ученым митохондрии преподнесли в 1979 г. До того времени считалось, что генетический код универсален и одни и те же триплеты кодируют одинаковые аминокислоты у бактерий, вирусов, грибов, растений и животных. Английский исследователь Беррел сопоставил структуру одного из митохондриальных генов теленка с последовательностью аминокислот в кодируемой этим геном субъединице цитохромоксидазы. Оказалось, что генетический код митохондрий крупного рогатого скота (как и человека) не просто отличается от универсального, он “идеален”, т.е. подчиняется следующему правилу: “если два кодона имеют два одинаковых нуклеотида, а третьи нуклеотиды принадлежат к одному классу (пуриновых — А, Г, или пиримидиновых — У, Ц), то они кодируют одну и ту же аминокислоту”. В универсальном коде есть два исключения из этого правила: триплет АУА кодирует изолейцин, а кодон АУГ — метионин, в то время как в идеальном коде митохондрий оба эти триплета кодируют метионин; триплет УГГ кодирует лишь триптофан, а триплет УГА — стоп-кодон. В универсальном коде оба отклонения касаются принципиальных моментов синтеза белка: кодон АУГ — инициирующий, а стоп-кодон УГА останавливает синтез полипептида. Идеальный код присущ не всем описанным митохондриям, но ни у одной из них нет универсального кода. Можно сказать, что митохондрии говорят на разных языках, но никогда — на языке ядра.
Как уже говорилось, в митохондриальном геноме позвоночных есть 22 гена тРНК. Каким же образом такой неполный набор обслуживает все 60 кодонов для аминокислот (в идеальном коде из 64 триплетов четыре стоп-кодона, в универсальном — три)? Дело в том, что при синтезе белка в митохондриях упрощены кодон-антикодонные взаимодействия — для узнавания используется два из трех нуклеотидов антикодона. Таким образом, одна тРНК узнает все четыре представителя кодонового семейства, отличающиеся только третьим нуклеотидом. Например, лейциновая тРНК с антикодоном ГАУ встает на рибосоме напротив кодонов ЦУУ, ЦУЦ, ЦУА и ЦУГ, обеспечивая безошибочное включение лейцина в полипептидную цепь. Два других лейциновых кодона УУА и УУГ узнаются тРНК с антикодоном ААУ. В целом, восемь разных молекул тРНК узнают восемь семейств по четыре кодона в каждом, и 14 тРНК узнают разные пары кодонов, каждая из которых шифрует одну аминокислоту.
Важно, что ферменты аминоацил-тРНК-синтетазы, ответственные за присоединение аминокислот к соответствующим тРНК митохондрий, кодируются в ядре клетки и синтезируются на рибосомах эндоплазматической сети. Таким образом, у позвоночных животных все белковые компоненты митохондриального синтеза полипептидов зашифрованы в ядре. При этом синтез белков в митохондриях не подавляется циклогексимидом, блокирующим работу эвкариотических рибосом, но чувствителен к антибиотикам эритромицину и хлорамфениколу, ингибирующим белковый синтез в бактериях. Этот факт служит одним из аргументов в пользу происхождения митохондрий из аэробных бактерий при симбиотическом образовании эвкариотических клеток [4].
Симбиотическая теория происхождения митохондрий
Гипотезу о происхождении митохондрий и растительных пластид из внутриклеточных бактерий-эндосимбионтов высказал Р.Альтман еще в 1890 г. За век бурного развития биохимии, цитологии, генетики и появившейся полвека назад молекулярной биологии гипотеза переросла в теорию, основанную на большом фактическом материале. Суть ее такова: с появлением фотосинтезирующих бактерий в атмосфере Земли накапливался кислород — побочный продукт их метаболизма. С ростом его концентрации усложнялась жизнь анаэробных гетеротрофов, и часть из них для получения энергии перешла от бескислородного брожения к окислительному фосфорилированию. Такие аэробные гетеротрофы могли с бOльшим КПД, чем анаэробные бактерии, расщеплять органические вещества, образующиеся в результате фотосинтеза. Часть свободно живущих аэробов была захвачена анаэробами, но не “переварена”, а сохранена в качестве энергетических станций, митохондрий. Не стоит рассматривать митохондрии как рабов, взятых в плен, чтобы снабжать молекулами АТФ не способные к дыханию клетки. Они скорее “существа”, еще в протерозое нашедшие для себя и своего потомства лучшее из убежищ, где можно затрачивать наименьшие усилия, не подвергаясь риску быть съеденными.
В пользу симбиотической теории говорят многочисленные факты:
— совпадают размеры и формы митохондрий и свободно живущих аэробных бактерий; те и другие содержат кольцевые молекулы ДНК, не связанные с гистонами (в отличие от линейных ядерных ДНК);Существует представление, что разные царства эвкариот имели разных предков и эндосимбиоз бактерий возникал на разных этапах эволюции живых организмов. Об этом же говорят отличия в строении митохондриальных геномов простейших, грибов, растений и высших животных. Но во всех случаях основная часть генов из промитохондрий попала в ядро, возможно, с помощью мобильных генетических элементов. При включении части генома одного из симбионтов в геном другого интеграция симбионтов становится необратимой.— по нуклеотидным последовательностям рибосомные и транспортные РНК митохондрий отличаются от ядерных, демонстрируя при этом удивительное сходство с аналогичными молекулами некоторых аэробных грамотрицательных эубактерий;
— митохондриальные РНК-полимеразы, хотя и кодируются в ядре клетки, ингибируются рифампицином, как и бактериальные, а эвкариотические РНК-полимеразы нечувствительны к этому антибиотику;
— белковый синтез в митохондриях и бактериях подавляется одними и теми же антибиотиками, не влияющими на рибосомы эвкариот;
— липидный состав внутренней мембраны митохондрий и бактериальной плазмалеммы сходен, но сильно отличается от такового наружной мембраны митохондрий, гомологичной другим мембранам эвкариотических клеток;
— кристы, образуемые внутренней митохондриальной мембраной, являются эволюционными аналогами мезосомных мембран многих прокариот;
— до сих пор сохранились организмы, имитирующие промежуточные формы на пути к образованию митохондрий из бактерий (примитивная амеба Pelomyxa не имеет митохондрий, но всегда содержит эндосимбиотические бактерии).
Новый геном может создавать метаболические пути, приводящие к образованию полезных продуктов, которые не могут быть синтезированы ни одним из партнеров по отдельности. Так, синтез стероидных гормонов клетками коры надпочечников представляет собой сложную цепь реакций, часть которых происходит в митохондриях, а часть — в эндоплазматической сети [5]. Захватив гены промитохондрий, ядро получило возможность надежно контролировать функции симбионта. В ядре кодируются все белки и синтез липидов наружной мембраны митохондрий, большинство белков матрикса и внутренней мембраны органелл. Самое главное, что ядро кодирует ферменты репликации, транскрипции и трансляции мтДНК, контролируя тем самым рост и размножение митохондрий. Скорость роста партнеров по симбиозу должна быть приблизительно одинаковой. Если хозяин будет расти быстрее, то с каждым его поколением число симбионтов, приходящихся на одну особь, будет уменьшаться, и, в конце концов, появятся потомки, не имеющие митохондрий. Мы знаем, что в каждой клетке организма, размножающегося половым путем, содержится много митохондрий, реплицирующих свои ДНК в промежутке между делениями хозяина. Это служит гарантией того, что каждая из дочерних клеток получит по крайней мере одну копию генома митохондрии.
Цитоплазматическая наследственность
Помимо кодирования ключевых компонентов дыхательной цепи и собственного белоксинтезирующего аппарата, митохондриальный геном в отдельных случаях участвует в формировании некоторых морфологических и физиологических признаков. К таким признакам относятся характерные для ряда видов высших растений синдром NCS (non-chromosomal stripe, нехромосомно кодируемая пятнистость листьев) и цитоплазматическая мужская стерильность (ЦМС), приводящая к нарушению нормального развития пыльцы. Проявление обоих признаков обусловлено изменениями в структуре мтДНК. При ЦМС наблюдаются перестройки геномов митохондрий в результате рекомбинационных событий, ведущих к делециям, дупликациям, инверсиям или инсерциям определенных нуклеотидных последовательностей или целых генов. Такие изменения могут вызывать не только повреждения имеющихся генов, но и появление новых работающих генов.
Цитоплазматическая наследственность, в отличие от ядерной, не подчиняется законам Менделя. Это связано с тем, что у высших животных и растений гаметы от разных полов содержат несопоставимые количества митохондрий. Так, в яйцеклетке мыши имеется 90 тыс. митохондрий, а в сперматозоиде — лишь четыре. Очевидно, что в оплодотворенной яйцеклетке митохондрии преимущественно или только от женской особи, т.е. наследование всех митохондриальных генов материнское. Генетический анализ цитоплазматической наследственности затруднен из-за ядерно-цитоплазматических взаимодействий. В случае цитоплазматической мужской стерильности мутантный митохондриальный геном взаимодействует с определенными генами ядра, рецессивные аллели которых необходимы для развития признака. Доминантные аллели этих генов как в гомо-, так и в гетерозиготном состоянии восстанавливают фертильность растений вне зависимости от состояния митохондриального генома.
Изучение геномов митохондрий, их эволюции, идущей по специфическим законам популяционной генетики, взаимоотношений между ядерными и митохондриальными генетическими системами, необходимо для понимания сложной иерархической организации эвкариотической клетки и организма в целом.
С определенными мутациями в митохондриальной ДНК или в ядерных генах, контролирующих работу митохондрий, связывают некоторые наследственные болезни и старение человека [6]. Накапливаются данные об участии дефектов мтДНК в канцерогенезе. Следовательно, митохондрии могут быть мишенью химиотерапии рака. Имеются факты о тесном взаимодействии ядерного и митохондриального геномов в развитии ряда патологий человека. Множественные делеции мтДНК обнаружены у больных с тяжелой мышечной слабостью, атаксией, глухотой, умственной отсталостью, наследующихся по аутосомно-доминантному типу. Установлен половой диморфизм в клинических проявлениях ишемической болезни сердца, что скорее всего обусловлено материнским эффектом — цитоплазматической наследственностью. Развитие генной терапии внушает надежду на исправление дефектов в геномах митохондрий в обозримом будущем.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Проект 01-04-48971.
Автор признателен аспиранту М.К.Иванову, создавшему рисунки к статье.Литература
1. Янковский Н.К., Боринская С.А. Наша история, записанная в ДНК // Природа. 2001. №6. С.10—18.
2. Минченко А.Г., Дударева Н.А. Митохондриальный геном. Новосибирск, 1990.
3. Гвоздев В.А. // Сорос. образоват. журн. 1999. №10. С.11—17.
4. Маргелис Л. Роль симбиоза в эволюции клетки. М., 1983.
5. Скулачев В.П. // Сорос. образоват. журн. 1998. №8. С.2—7.
6. Игамбердиев А.У. // Сорос. образоват. журн. 2000. №1. С.32—36.