№ 7, 2003 г.
© Л.В.Полищук
СКОРОСТЬ РАЗМНОЖЕНИЯ
И УГРОЗА ВЫМИРАНИЯ ВИДАЛ.В. Полищук
Леонард Владимирович Полищук, к.б.н., научн. сотр. кафедры общей экологии биофака МГУ
В наши дни скорость вымирания видов на два-три порядка выше той, что была до палеолита [1]. Только c начала XVII в. из примерно 4 тыс. видов млекопитающих по крайней мере 63 - уже нет. Среди наиболее известных потерь отечественной фауны - предок европейского крупного рогатого скота тур (Bos primigenius), родоначальник домашней лошади тарпан (Equus gmelini), стеллерова корова (Hydrodamalis gigas), которая была истреблена человеком спустя всего 27 лет после ее открытия. Однако причины вымирания растений и животных не всегда столь очевидны. В современном мире это нередко происходит в результате сокращения пригодных местообитаний, а также вселения хищников или паразитов, которым аборигенные виды не могут противостоять.
В середине XX в. Международный союз охраны природы и природных ресурсов (МСОП) начал сбор информации о редких и находящихся под угрозой исчезновения видах, что привело в дальнейшем к созданию Красных книг. Критерии включения вида в “красные списки”, как теперь их еще называют, со временем совершенствовались и детализировались, но основными были и остаются: значительное сокращение численности и/или ареала вида (теперь обычно считается, что на протяжении не менее трех поколений) или же его редкость (низкая общая численность, узкий ареал). Однако, как бы тщательно ни собирались эти сведения и какими квалифицированными ни были специалисты, занимающиеся этим делом, избавиться от субъективности при формировании “краснокнижных” списков очень трудно [2]. Поэтому для решения, какой вид действительно нуждается в повышенном внимании, важно найти некие дополнительные критерии, основанные на строго измеряемых характеристиках вида, по которым можно было бы судить о его уязвимости. Именно это я и попытался сделать - оценить уязвимость вида, используя в качестве материала данные по демографии и массе тела млекопитающих [3, 4].
Вероятность вымирания и задача о разорении игрока
Вымирание грозит, по-видимому, в основном медленно размножающимся видам. Наглядные тому примеры - стеллерова корова, производившая на свет лишь одного детеныша раз в два года, или птица додо (Raphus cucullatus) из семейства дронтовых, в кладке которой было только одно яйцо. В исчезновении таких видов принято винить человека, и это, конечно, верно, но ведь зайцев тоже истребляли, тем не менее им, как и другим видам из отряда зайцеобразных (кроликам, пищухам), вымирание не грозит. Более того, это единственный отряд из фауны млекопитающих России и сопредельных регионов, ни один представитель которого не внесен в Красную книгу, несмотря на то, что зайцы всегда были излюбленным объектом охоты, а ареал кроликов занимает совсем небольшое пространство на юге Украины и в Молдавии. В чем же тут дело? Их, скорее всего, спасает баснословная, вошедшая в поговорку скорость размножения. Связь между вероятностью вымирания вида и скоростью его размножения можно установить, обратившись к известной из теории вероятностей задаче о разорении игрока [5].
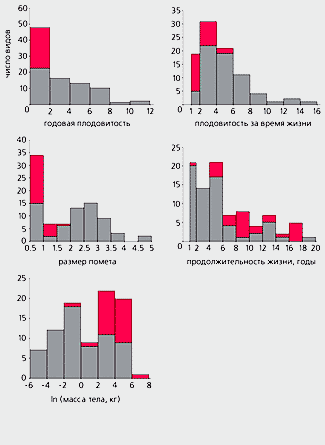
Распределение краснокнижных (выделено цветом) и некраснокнижных видов млекопитающих фауны бывшего Советского Союза по плодовитости за год и за время жизни, размеру помета (эти три признака выражены в числе дочерей), продолжительности жизни взрослых особей и массе тела. По условию задачи, игра, в которой один из участников ставит на кон z рублей (чтобы задача не выглядела слишком искусственной, допустим, что в другой раз второй игрок поставит эту же сумму), состоит в проведении ряда испытаний, причем для владельца начального капитала вероятность выиграть рубль и добавить его к имеющейся сумме при каждом испытании равна p, а проиграть рубль - q (p>q, в противном случае проигрыш практически гарантирован). Для выигрыша надо накопить сумму, много большую начального капитала; проигрыш - это потеря z рублей, т.е. разорение. Какова же вероятность разорения (P)? Решение задачи: P=(p/q)–z.
Математически процесс вымирания особей можно описать точно так же, как потерю денег, только вместо вероятности выигрыша p фигурирует величина, пропорциональная удельной (в расчете на особь) рождаемости b, вместо вероятности проигрыша q - величина, пропорциональная удельной смертности d, а роль начального капитала играет число особей n, с которым ассоциируется вероятность вымирания. В итоге вероятность вымирания вида (популяции) описывается следующей моделью: P=(b/d)–n, а соответствующую теорию можно назвать демографической теорией вымирания [6].
Биологическая интерпретация модели вымирания, основанной на задаче о разорении игрока, очень проста. Поскольку отношение b/d - это количество потомков, которое самка производит за время жизни (так как 1/d - это средняя продолжительность жизни), модель показывает, что вероятность вымирания тем меньше, чем больше скорость размножения. Это вполне согласуется с общепринятыми представлениями экологов. Однако использовать эту модель в качестве количественной теории довольно сложно. Во-первых, ее невозможно “параметризовать”, т.е. оценить значение n, поскольку в нашем распоряжении нет независимых оценок P. Во-вторых, по этой же причине ее трудно верифицировать, т.е. сделать прогноз вероятности вымирания и сравнить его с реальностью. Наконец, как и большинство математических построений, опирающихся не столько на конкретные биологические знания, сколько на здравый смысл, модель представляет собой слишком жесткую конструкцию для описания реальности. Так, в ней не учитывается возможная связь вероятности вымирания с другими характеристиками вида, например, с массой тела. Между тем на этой зависимости основан известный закон увеличения размеров организмов в филогенетической ветви (или так называемое правило Депере), согласно которому “по мере хода геологического времени выживающие формы увеличивают свои размеры (а следовательно, вес) и затем вымирают” (В.И.Вернадский). Вполне может быть, что и другие показатели связаны с вероятностью вымирания нисколько не хуже, чем отношение b/d. Модель, претендующая на описание реальности, должна быть гибкой, и самое главное для ее построения необходимы количественные оценки вероятности вымирания или какого-либо иного показателя уязвимости вида.
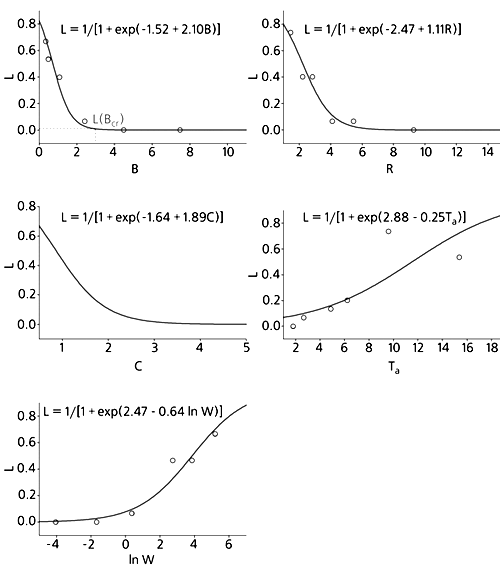
Вероятность попадания вида в Красную книгу (L), построенная по логистической регрессии, в зависимости от плодовитости за год (B) и за время жизни (R), размера помета (C), продолжительности жизни взрослых особей (Ta) и массы тела (W). Уравнения и соответствующие им кривые построены по всему массиву данных (т.е. по 90 точкам, каждая из которых соответствует отдельному виду). На четырех графиках из пяти нанесены точки, соответствующие эмпирической частоте попадания в Красную книгу, которая получается при разбиении упорядоченного по данному признаку набора из 90 видов на определенное число групп (здесь на 6 по 15 видов в каждой) и подсчете доли краснокнижных видов в каждой группе. Упорядоченный ряд нельзя построить только для размера помета, поскольку у многих видов размер помета одинаков. На графике зависимости от годовой плодовитости показана пороговая плодовитость Bcr = 2.9 дочерей/год, соответствующая вероятности попасть в Красную книгу L(Bcr) = 0.01.Красная книга и скорость размноженияВ большинстве случаев реально оценить вероятность вымирания невозможно, к тому же она может оказаться столь высокой (например, при очень низкой численности), что уже поздно принимать меры по спасению вида. Поэтому гораздо важнее оценить перспективы вида, т.е. вероятность угрозы его вымирания. Как известно, виды, которым уже реально угрожает вымирание, занесены в списки Красной книги. Учитывая эту информацию, можно попробовать оценить вероятность угрозы вымирания (иными словами - вероятность стать кандидатом в Красную книгу) и для любого другого вида. Такие оценки могут быть полезны как для развития теории, так и для практики, поскольку позволяют заранее, до наступления реальной опасности, выявлять виды, нуждающиеся в защите.
В качестве объекта были взяты млекопитающие - одна из наиболее хорошо изученных групп животных. Общей сводки по демографии млекопитающих обнаружить не удалось, поэтому из многочисленных, в основном отечественных, публикаций (использовано более 130 литературных источников) были собраны сведения о 90 видах, т.е. примерно 30% от всех млекопитающих фауны бывшего СССР. “Краснокнижными” в нашей выборке оказались 25 видов, или 28%, притом что во всей фауне их доля составляет примерно 25% (примерно из 300 видов млекопитающих СССР в “Красной книге СССР” представлено 77, не считая подвидов) [7].
Для каждого выбранного вида были найдены четыре демографические характеристики: средние величины плодовитости за год и за время жизни (выраженные в количестве дочерей, производимых самкой за год и за среднее время жизни взрослых особей соответственно), число дочерей в помете и средняя продолжительность жизни взрослых особей (т.е. после достижения половой зрелости). Помимо этого был использован еще один признак - средняя масса тела взрослых самок.
Дабы не вызывать лишних вопросов у читателя, сразу оговорюсь: принимались во внимание только те характеристики, которые были получены в дикой природе, а не в условиях зоопарка или вивария. То, что в расчетах учитывалось лишь число самок, - дань установившейся в демографии традиции (число самок получалось из общего числа детенышей делением пополам). Кроме того, замечу, что использованные демографические характеристики относятся к виду в целом, а в Красной книге многие виды представлены на уровне отдельных географических подвидов; однако, как показала специальная проверка, характеристики “краснокнижных” подвидов вполне согласуются с усредненными видовыми показателями.
Все 90 видов были расположены в порядке убывания значений по каждому из пяти изученных признаков, и достаточно было простого просмотра списков, чтобы заметить в них крайне неравномерное распределение “краснокнижных” видов [3]. Из-за недостатка места невозможно привести здесь данные для всех 90 видов по всем признакам, ограничимся для примера лишь одним из них - плодовитостью за время жизни. Разобьем упорядоченный по этому признаку список на девять групп по 10 видов и посмотрим, сколько в каждой из них охраняемых. Среди наиболее плодовитых видов нет ни одного “краснокнижного”, а среди наименее - их восемь (табл.1). Более того, по мере убывания плодовитости количество таких видов в группах закономерно возрастает или остается тем же, но никогда не уменьшается.
Столь статистически не случайное распределение охраняемых видов наблюдается при рассмотрении всех изученных признаков: в Красную книгу попадают, как правило, крупные и долгоживущие млекопитающие, с небольшим числом детенышей в помете и низкой скразмножения. Так, годовая плодовитость “краснокнижных” видов в среднем более чем в шесть раз ниже, чем неохраняемых (0.56±0.07 против 3.56±0.34 дочерей в год соответственно), а их плодовитость за время жизни ниже более чем в два раза (2.20±0.19 против 5.02±0.37 дочерей за жизнь).
Полученные результаты находятся в явном соответствии с теорией вымирания, согласно которой большему риску вымирания подвержены медленно, а не быстро размножающиеся виды [6, 8]. Это значит, что вероятность оказаться под угрозой вымирания, или вероятность попадания вида в Красную книгу, - столь же объективный показатель уязвимости вида, как и более традиционная, но труднее определяемая практически, вероятность вымирания.
Вероятность попадания в Красную книгу и логистическая регрессия
Примерно оценить вероятность попадания в Красную книгу можно непосредственно по таблице, в которой виды упорядочены по определенному признаку. Так, если среди десяти наименее плодовитых видов восемь находятся в Красной книге, то для всех видов с плодовитостью из этого диапазона вероятность оказаться там равна примерно 0.8. Однако такая оценка не вполне нас удовлетворяет хотя бы потому, что ранжирование видов не всегда можно провести однозначно (поскольку разные виды могут иметь одинаковое значение признака), да и сам список можно разбить на группы многими разными способами.
Попробуем рассуждать иначе. Допустим, нас интересует судьба некоего вида, биология которого хорошо изучена - известны скорость размножения, продолжительность жизни и т.д. Естественно, он находится под влиянием факторов среды, в том числе и тех, которые могут угрожать его существованию. Будущее вида может сложиться двояко: его численность, несмотря на воздействие неблагоприятных факторов, может оставаться стабильной или даже расти (например, благодаря высокой рождаемости), а может от поколения к поколению сокращаться, что в результате будет замечено, и вид окажется на страницах Красной книги. На математическом языке это можно описать так: изначально вид имел определенную вероятность попасть в Красную книгу, а то, что произошло с ним в итоге, - единичная реализация вероятностного процесса. Задача состоит в том, чтобы эту вероятность найти. Классическое частотное определение вероятности, как, например, при бросании монеты, применить в данном случае нельзя просто потому, что не бывает одинаковых видов и в реальности наш мысленный эксперимент нельзя повторить. Но ведь каждый вид характеризуется определенным признаком (и даже не одним, а несколькими), который может быть связан с интересующей нас вероятностью монотонной зависимостью; к тому же, как мы помним, в нашем распоряжении множество (выборка) видов.
Описанный подход был использован для построения зависимостей вероятности попадания вида в Красную книгу от каждого из рассматриваемых демографических признаков и массы тела по выборке из 90 видов. Качественно результат не отличался от полученного ранее при простом ранжировании видов: более высокую вероятность попасть в Красную книгу имеют крупные и медленно размножающиеся виды. Теперь, однако, в наших руках оказалась количественная модель (регрессионная зависимость), позволяющая прогнозировать интересующую нас вероятность.
Следующий шаг статистического анализа - построение зависимости вероятности от комбинаций признаков (множественных регрессионных моделей) - не привел к существенному улучшению качества модели. И это понятно, ведь все признаки взаимосвязаны: например, особи мелких видов живут недолго и, как правило, размножаются с большой скоростью. Поэтому в дальнейшем будем рассматривать только однофакторные модели. Но прежде чем перейти к детальному изучению полученных зависимостей, стоит определить, какая из них точнее соответствует реальности, и именно ей уделить основное внимание.
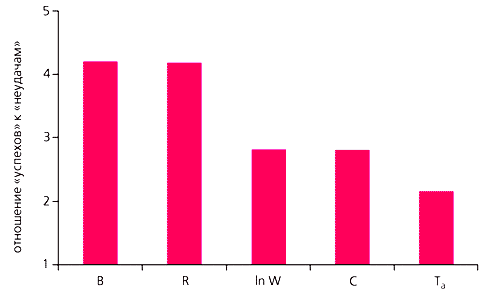
По каждой зависимости рассчитали вероятность попадания в Красную книгу для каждого конкретного вида, исходя из значения характеризующего его признака. Если эта вероятность была больше 0.5, то полагали, что вид находится под угрозой и должен присутствовать в Красной книге, если меньше - что нет. Результат такой обратной классификации сравнивали с реальностью, т.е. присутствует или отсутствует данный вид в Красной книге. Проведя сравнение по всем 90 видам, получали таблицу сопряженности 2ґ2, в двух клетках которой указано число правильно классифицированных видов, а в двух других – неправильно. Произведение чисел, стоящих в двух первых клетках, деленное на произведение чисел в двух последних, называется в теории логистической регрессии отношением шансов. Среднее соотношение между числом правильных (“успех”) и неправильных (“неудача”) классификаций находили как корень квадратный из отношения шансов. Оказалось, что наилучшее отношение “успехов” к “неудачам” обеспечивают два признака - плодовитость за год и за время жизни. Именно эти признаки непосредственно связаны со скоростью размножения. Поскольку точно определить продолжительность жизни взрослых особей в природе довольно сложно, наибольшую практическую ценность представляет вероятность, рассчитанная по годовой плодовитости.
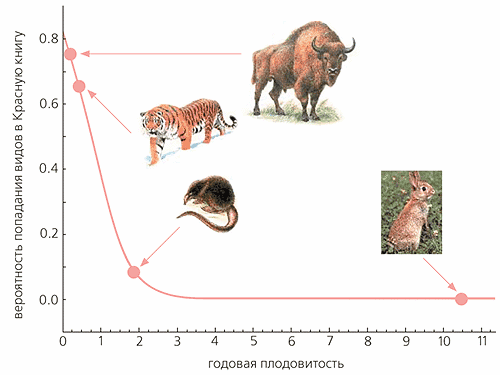
Вероятность попадания вида в Красную книгу находится в нелинейной зависимости от годовой плодовитости, причем эта зависимость имеет протяженное, практически горизонтальное плато в области нулевых значений вероятности. Так, при увеличении плодовитости до 2.9 дочерей в год вероятность попадания вида в Красную книгу уменьшается до 0.01, а для еще более быстро размножающихся видов становится почти нулевой. Если пренебречь столь малыми величинами, то указанное значение годовой плодовитости можно считать пороговым. (Видов с годовой плодовитостью больше 2.9 в нашей выборке из 90 оказалось 30, что совсем немало.) Подчеркнем еще, что принятое пороговое значение не является артефактом модели, так как в выборке действительно нет ни одного “краснокнижного” вида с годовой плодовитостью больше 1.75 дочерей (эта величина характеризует выхухоль). Таким образом, 2.9 дочерей в год - пороговая плодовитость с определенным “запасом прочности”.
Для области высоких значений вероятности характерно то, что даже при самой низкой плодовитости вероятность попадания вида в Красную книгу равна 0.8, а отнюдь не 1. Это значит, что в выборке нет ни одного вида, безусловно обреченного на такое сокращение численности, которое чревато вымиранием.
Понятно, что этот оптимистичный прогноз верен только при сохранении существующих условий. Любой вид можно довести до гибели, если приложить к этому специальные усилия или в случае глобальной катастрофы. Тем не менее даже с этой оговоркой прогноз, основанный на скорости размножения, дает определенное представление о том, какие виды более, а какие менее подвержены риску вымирания. Чем быстрее размножается вид, тем, как правило, труднее снизить его численность до такой степени, чтобы поставить его существование под угрозу.
Какие виды нуждаются в охране в первую очередь
Вероятность попадания вида в Красную книгу опирается лишь на “внутренние” характеристики вида (такие как средние, типичные для него годовая плодовитость, размер помета и т.д.) и не учитывает внешние условия, которые между тем постоянно меняются и влияют на эти характеристики. Тем не менее такой подход может весьма эффективно описывать реальность. Вопрос только в том, к какой реальности он приложим, вернее, каковы границы его применимости.
Наш подход относится к сфере макроэкологии (термин предложен Дж.Брауном и Б.Маурером в 1989 г. [9]). Эта часть экологии изучает явления, происходящие на больших пространствах, когда биологические, в частности демографические, свойства видов выходят на первый план, а изменчивость среды, доставляющая в других ситуациях массу неприятностей экологам, отступает на второй. На языке вероятностной теории вымирания это означает, что в макромасштабе основное значение имеет демографическая стохастичность и гораздо меньшее - средовая. Почему так происходит? Демографическая стохастичность зависит от скорости роста численности (именно с ней мы имеем дело в этой работе). Средовую стохастичность можно описать так: в последовательные отрезки времени среда становится то лучше, то хуже, и, если эти отклонения велики, на каком-то отрезке среда может стать совсем неблагоприятной, что приведет к вымиранию вида. Говоря формально, средовая стохастичность зависит от дисперсии скорости роста численности. Если дисперсия велика, то даже при большой скорости роста вероятность вымирания все равно велика. Однако на большом пространстве эта дисперсия стремится к нулю, потому что обычно отдельные локальные популяции в разных точках пространства могут испытывать противоположно направленные средовые воздействия, взаимно уравновешивающие друг друга. Здесь работает закон больших чисел, где “большое число” - количество удаленных друг от друга, а потому почти независимых популяций. Таким образом, на большом пространстве эффект средовой неопределенности относительно невелик. Однако при обычном, среднем уровне численности эффект демографической стохастичности также невелик, поскольку он проявляется только в случае низкой численности.
Иная ситуация складывается в случае, когда из-за неумеренного промысла или природных катаклизмов (неурожая кормов, эпизоотии, холодной или многоснежной зимы и т.д.) численность всех локальных популяций и вида в целом одновременно сокращается. Такие катастрофы довольно редки, однако теоретически любой вид может оказаться в подобной ситуации. Тогда, по-видимому, и решается, кому суждено погибнуть или в лучшем случае попасть в Красную книгу, а кому - нет. В периоды низкой численности эффект средовой стохастичности на большой территории по-прежнему невелик, и судьба вида оказывается полностью зависимой от демографической стохастичности. Виды с высокой рождаемостью (самки которых, по нашей оценке, приносят более 2.9 дочерей в год) быстро возвращаются к прежнему, нормальному уровню численности, им вымирание, как правило, не грозит. Напротив, медленно размножающиеся виды задерживаются в состоянии низкой численности на длительное время, поскольку им труднее компенсировать последствия, например увеличения промысла.
Яркая иллюстрация тому - катастрофическая ситуация, сложившаяся в последнее время с популяцией сайгака (Saiga tatarica). Замечу, в 20-х годах прошлого столетия, когда Красных книг еще не было, в результате неконтролируемого промысла численность и ареал этого млекопитающего резко сократились. В 30-е годы в связи с прекращением контрабандного вывоза сайгачьих рогов в Китай и во время Великой Отечественной войны, когда людям было не до сайгака, его численность постепенно начала восстанавливаться. А в 50-е годы стало ясно, что виду ничто не угрожает. Теперь ситуация, похоже, повторяется. Рога сайгака всегда ценились в китайской медицине, а сейчас их стали еще и рекламировать в качестве альтернативы рогу носорога. В результате на китайском рынке резко вырос спрос на рога сайгака, и браконьеры начали истреблять их (прежде всего самцов), что поставило под угрозу существование всей популяции. А ведь до октября 2002 г., когда сайгак был внесен в Красную книгу МСОП, он вообще не числился среди “краснокнижных” видов. Хотя плодовитость сайгака, равная 0.75 дочерей в год, выше, чем у многих других копытных, соответствующая ей вероятность попадания в Красную книгу составляет примерно 0.5. Это совсем не маленькая вероятность: она означает, что каждый второй вид с такой годовой плодовитостью оказывается под угрозой вымирания. Ясно, что даже в периоды высокой численности сайгак нуждается в пристальном внимании экологов и специалистов по охране природы.
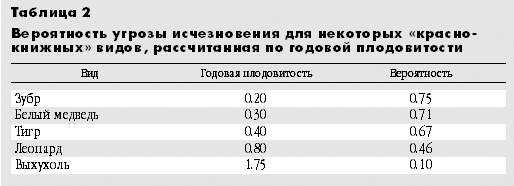
Новый подход позволяет заблаговременно выявлять уязвимые виды и распределять природоохранные ресурсы между ними пропорционально их вероятности попадания в Красную книгу, а не по принципу “всем сестрам по серьгам”. При этом затраты на мониторинг видов, которые находятся в относительной безопасности (т.е. с годовой плодовитостью больше порогового значения - 2.9 дочерей в год), могут составлять некий базовый минимум и быть примерно одинаковыми для разных видов независимо от их плодовитости. На те виды, годовая плодовитость которых меньше порогового значения, напротив, следует тратить больше средств, и они должны распределяться в примерном соответствии с вероятностью их попадания в Красную книгу. Так, вероятность попасть в Красную книгу для белого медведя и амурского тигра примерно одинакова, для леопарда она в полтора раза меньше, а для выхухоли - реликтового вида и эндемика России - в семь раз меньше (табл.2). Можно полагать, что и ресурсов на мониторинг и охрану леопарда и выхухоли по сравнению с белым медведем или тигром должно выделяться примерно во столько же раз меньше. А что происходит на самом деле?
Согласно федеральной целевой программе Российской Федерации на 2002-2010 гг., на мониторинг амурского тигра и белого медведя выделяется 527.1 и 431.4 млн руб. соответственно [11]. Большой объем и примерное равенство выделяемых средств вполне согласуются с высокой и приблизительно одинаковой вероятностью попадания этих видов в Красную книгу. Однако доля этих средств в общем объеме затрат далеко не соответствует соотношению между вероятностями: на эти виды приходится 95% ресурсов, выделяемых на мониторинг всех животных и растений Российской Федерации. Если учесть, что в Красной книге России только млекопитающих 65 видов (без учета подвидов) и что многим из них угрожает реальная опасность (амурского леопарда осталось всего 20-30 особей [12], а численность выхухоли за последние 30 лет снизилась в два раза [13]), становится понятным, что имеющихся природоохранных ресурсов катастрофически не хватает.
Я далек от мысли, что вероятность оказаться под угрозой вымирания, рассчитанная по скорости размножения вида, может служить единственным критерием для распределения природоохранных ресурсов. Понятно, например, что организация экспедиции на Крайний Север для изучения состояния белого медведя стоит намного дороже, чем аналогичное обследование выхухоли в Центральной России. Тем не менее распределение ресурсов, учитывающее только экономические и политические соображения и не принимающее во внимание биологические характеристики животных, вряд ли может быть эффективным с точки зрения сохранения биоразнообразия. Непосредственное практическое следствие из предлагаемого здесь подхода может состоять в том, чтобы, исходя из соотношения вероятностей попадания в Красную книгу и взяв за основу, например, затраты на охрану амурского тигра как одного из наиболее благополучных с точки зрения привлечения общественного внимания видов, оценить, хотя бы примерно, сколько недодано другим находящимся под угрозой видам. Такие оценки могли бы служить основанием для привлечения дополнительных средств для охраны животных, - причем именно тех, которым в настоящее время уделяется неоправданно мало внимания.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Проект 01-04-48384.
Я чрезвычайно благодарен А.М.Гилярову, С.Пимму (S.Pimm), И.Хански (I.Hanski), Ю.Ю.Дгебуадзе, В.В.Малахову, Н.А.Формозову, В.Б.Цейтлину, Б.И.Шефтелю, Ю.Шига (Y.Shiga), а также многим другим, способствовавшим выполнению этой работы.Литература 1. Pimm S.L., Russell G.J., Gittleman J.L. et al. // Science. 1995. V.269. P.347-350.
2. Гиляров А. // Наука и жизнь. 1976. №12. С.132-139.
3. Polishchuk L.V. // Журн. общ. биологии. 2002. Т.63. ?2. С.99-111.
4. Polishchuk L.V. // Science. 2002. V.297. P.1123.
5. Феллер В. Введение в теорию вероятностей и ее приложения. Т.1. М., 1967.
6. MacArthur R.H. Geographical ecology. N.Y., 1972.
7. Млекопитающие // Красная книга СССР. Т.1. М., 1984. С.9-97.
8. Southwood T.R.E. // Theoretical ecology: principles and applications. Oxford, 1976. P.26-48.
9. Brown J.H., Maurer B.A. // Science. 1989. V.243. P.1145-1150.
10. Новиков Б.В. Росомаха. М., 1993.
11. Подпрограмма “Сохранение редких и исчезающих видов животных и растений” // Федеральная целевая программа “Экология и природные ресурсы России (2002-2010 годы)”. М., 2001. (http://www.priroda.ru/bibl/knigi/1748_0860_2001.pdf)
12. Млекопитающие // Красная книга Российской Федерации. М., 2001. С.599-750.
13. http://www.biodiversity.ru/programs/vyhuhol.html (сайт Центра охраны дикой природы)
Июнь 2003