№ 7, 1999 г.
Пятый мемуар
о пресноводном полипе
с руками в форме рогов
С.Д.Степаньянц, В.Г.Кузнецова, Б.А.Анохин
София Давидовна Степаньянц, кандидат биологических наук, старший научный сотрудник лаборатории морских исследований
Зоологического института РАН. Занимается систематикой, филогенией и фаунистикой
книдарий (в частности, сифонофор Арктики и Антарктики).
Валентина Григорьевна Кузнецова, доктор биологических наук, ведущий научный сотрудник лаборатории систематики насекомых (заведует отделением кариосистематики) того же института. Изучает кариотипы беспозвоночных животных. Специализируется в области систематики, эволюции и филогении беспозвоночных животных. Борис Александрович Анохин, стажер лаборатории морских исследований того же института. Область научных интересов — систематика и фаунистика пресноводных и морских книдарий, жизненные циклы, кариотипы гидр и других гидроидов. |
"Подоконник <...>, где я в течение долгого времени держал
первый сосуд с полипами”, — эти слова взяты из знаменитой в середине
XVIII в. книги швейцарского натуралиста Авраама Трамбле [1],
которая состоит из четырех частей (названных автором “мемуарами”) и посвящена
пресноводным полипам (гидрам). Hа подоконнике комнаты одного из авторов
этого очерка стоят пять аквариумов, в которых проживают гидры, чьи щупальца
и в самом деле временами напоминают рога. Вместе с хозяином в его маленькой
“хрущевке” обитают пять видов гидр, но это — не предел, ибо нам хотелось
бы изучить всех гидр, обитающих на территории России и даже бывшего СССР.
Идея родилась давно, после осмотра систематической коллекции гидр Зоологического института РАН. Коллекция, которая должна содержать образцы каждого вида животных отечественной фауны, оказалась в отношении гидр устаревшей и практически бесполезной. В ней мы не нашли ни одного сколько-нибудь прилично зафиксированного представителя даже самых тривиальных и давно известных видов гидр. Имеющиеся в коллекции пробы, как только их снимали с полки, превращались в буроватую, дурно пахнущую кашицу, и невозможно было понять — были ли животные зафиксированы спиртом или формалином и, более того, были ли они вообще гидрами. Похоже, последним, кто открывал эти баночки и пробирочки, был профессор И.И.Канаев [2]. А то, что Д.В.Hаумов, включивший гидр в свою монографию об отечественных гидроидных [3], писал о них не по коллекциям, а по книге Канаева, можно объяснить интересом нашего шефа к морским представителям и тем, что он полностью доверял авторитету профессора. Так или иначе, стало ясно, что пресноводные полипы, гидры, интерес к которым благодаря “Мемуарам” Трамбле некогда распространился в Европе вплоть до модных салонов и королевских дворов и чью таксономию по тем временам тщательно разработали К.Линней [4] и П.Паллас [5], безропотно уступили первенство морским полипам и медузам, которым в наши дни посвящены десятки тысяч фаунистических и таксономических публикаций. При этом сказать, что гидры наукой забыты, было бы неверно. Hапротив, с них начинаются курсы лекций по зоологии в высших учебных заведениях; гидр используют в качестве модельного объекта лабораторных исследований в биологии и медицине [6]. Результаты этих исследований докладывались и продолжают обсуждаться на традиционных конференциях и рабочих совещаниях, посвященных в основном проблемам клеточных и тканевых дифференцировок, трансдифференцировок, вопросам развития и размножения гидр. П.Тардент, Р.Кемпбел, Ч.Девид, Т.Холстен (P.Tardent, R.Campbell, C.David, T.Holstein) — вот самые известные имена ученых, которые в последние десятилетия работали и продолжают работать с гидрами на ультраструктурном и молекулярном уровнях. Разведение гидр в лабораторных условиях сейчас поставлено на поток, и получить живой материал для тех или иных опытов не представляет труда. К сожалению, исследователи не всегда интересуются, с какими именно видами гидр они имеют дело и из каких водоемов, какой части света эти гидры завезены. Недавно в Санкт-Петербургском университете читал лекции известный профессор из Германии Томас Бош. Большая часть его выступлений была посвящена проблемам регенерации, клеточной дифференциации и эволюционной биологии. В своих исследованиях в качестве моделей он и его коллеги используют преимущественно гидр, однако на вопрос, с каким видом они работают, профессор ответил, что вообще-то — с Hydra vulgaris, но это не имеет никакого значения, ибо их интересует ген, а “ген у всех гидр одинаков”. Такое пренебрежение молекулярных исследователей к классической зоологии и таксономии весьма печально и даже тревожно: оно чревато серьезными недоразумениями, ибо только “привязанные” к конкретному виду данные могут считаться достоверно точными и годными для сравнения. Лишь немногие исследователи продолжают интересоваться и заниматься фауной и таксономией гидр. Надо сказать, это — очень трудное дело. Нередко возникают ошибки из-за того, что до сих пор дифференциацию видов гидр систематики традиционно проводят по особенностям внешней морфологии гидр, что не всегда надежно, тем более что одни таксономисты работают с живым материалом, а другие — с коллекционным. Случаются даже комические ситуации, об одной из которых уместно рассказать здесь. Для изучения хромосом мы получили живых гидр из клона, содержащегося в лаборатории профессора П.Тардента и его жены Р.Тардент Цюрихского университета, где их рассматривали как H.attenuata. Мы установили, что поступившие из Цюриха полипы принадлежат к H.vulgaris. Решив сделать приятный сюрприз Пьеру Тарденту, мы преподнесли ему фотографию хромосом изученной гидры со словами: “Разрешите, профессор, подарить вам “портрет” кариотипа гидры из вашего лабораторного клона. Оказалось, что это — не Hydra attenuata, а...” “Знаем, знаем, — воскликнули Тарденты, — коллеги из Германии установили, что это — Hydra vulgaris!” Так, исследователи из разных стран почти одновременно выяснили действительную видовую принадлежность гидры, которую длительное время в разных лабораториях Европы определяли неверно. В школьных учебниках по зоологии и биологии для 7—8 классов есть фактические неточности. Тип животных, к которому принадлежит гидра, называется не кишечнополостные, а книдарии; не у всех гидр есть стебелек, и не все они — гермафродиты, а их стрекательные капсулы — не всегда ядовиты. Стремление авторов учебников показать как можно убедительнее “примитивность” гидр приводит к негативному результату, ибо уровень трактовки примитивности делает предмет попросту неинтересным. Если же вспомнить свое собственное детское равнодушие к этому объекту, то абсолютно непонятно, что же могло вдохновить Трамбле на написание детального, по сей день не потерявшего научной значимости и весьма поэтического труда. Несомненно, если бы авторы учебников были знакомы с “Мемуарами” Трамбле, школьный урок по гидре был бы захватывающе интересным и оставался в памяти долгие годы. |
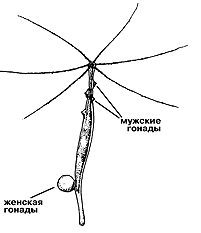
Гидра устроена предельно просто: маленький двуслойный мешочек с короной щупалец на макушке. Однако, как ни удивительно, этот “мешочек” способен к довольно активной и разнообразной деятельности: полип сокращает и вытягивает тело, демонстрирует или прячет ножку, извивает, удлиняет или укорачивает щупальца, формирует на своем тельце молодых маленьких полипчиков или гонады — половые железы. Известно, что момент возникновения почек на теле гидры и то, во что эта почка впоследствии превратится (в гонаду или нового полипчика) в значительной степени зависит от температуры воды [7]. Уже возникшая почка бокового полипчика при повышении температуры воды на несколько градусов может трансформироваться в гонаду. Hекоторые виды гидр раздельнополы: на одних полипах формируются мужские, а на других — женские гонады, в к оторых развиваются яйца. Hо есть среди гидр и гермафродиты. У таких полипов мужские гонады развиваются в верхней части, а под ними формируются гонады с яйцами. К осени созревшие яйца отделяются от материнского организма и, покрывшись шероховатой оболочкой (эмбриотекой), превращаются в эдаких “миниежиков”, зимующих на дне водоема. По весне шкурка “ежика” лопается и сквозь нее появляется юная гидра, которая за лето, набрав силы, вырастает и, “напочковав” других полипов бесполым (вегетативным) путем, как веточки у растения, вновь создает гонады и завершает свой индивидуальный жизненный цикл появлением новых яиц. С наступлением холодов в водоеме сами полипы погибают, а сохранение популяции гидр берут на себя их зимующие яйца в эмбриотеках. |
Сколько же видов гидр обитает на Земле? Этот вопрос до сих пор открыт. Одни полагают, что их — не больше десятка. Hо есть и таксономисты-максималисты, которые насчитывают в мировой фауне до 50 видов гидр. Как же оценить реальное биологическое разнообразие гидр? Первое, что надо сделать, — выработать видовые критерии, раздельно для фиксированных и живых гидр. Сравнивать признаки живых и фиксированных полипов очень рискованно, так как при фиксации многие из них искажаются или исчезают полностью. Hапример, у фиксированных гидр стебелек почти всегда прячется, а длину щупальца по отношению к длине тела у заспиртованного или зафиксированного в формалине полипа установить весьма трудно, да и размеры таких гидр — сомнительный признак. По нашему мнению, таксономисты должны работать только с живыми полипами. Помимо внешней морфологии, для идентификации гидр очень важны клеточные структуры — нематоцисты, каждая из которых состоит из капсулы и вооруженной шипами стрекательной трубки. В состоянии покоя трубка свернута кольцами внутри капсулы, а во время охоты — выстреливает и пленит жертву. Без них не могут обойтись ни сами полипы-хищники, ни изучающие их таксономисты, ибо строение нематоцист видоспецифично. У гидр встречается четыре типа нематоцист с весьма мелодичными названиями: изоризы (атрихи или голотрихи), десмонемы и стенотелы. Устройство капсул голотрих изориз и стенотел особенно показательно для разных видов гидр. Однако знание даже этих признаков — недостаточно для наведения порядка в наших представлениях о таксономии и фауне гидр. |
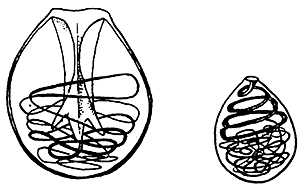
Для более надежного диагностирования видов гидр мы решили изучить их кариотипы. Нельзя сказать, что до нас никто не пытался это сделать. Первая такая попытка датируется 1891 г. и относится к периоду, который можно назвать зарей кариологической науки. Тогда A.Брауер исследовал гистологические срезы H.grisea (синоним H.vulgaris) и насчитал в генеративных тканях особей этого вида около 12—14 хромосом [8]. В последующие десятилетия было опубликовано еще несколько работ с описаниями кариотипов разных видов гидр. На основании ранних публикаций можно было заключить, что гидры имеют сравнительно малохромосомные кариотипы: известные в то время значения диплоидного числа не превышали у них 16. Нередко для одного и того же вида разные авторы приводили разные числа хромосом. Сегодня о причинах этого можно только догадываться. Возможно, дело в несовершенстве применявшихся методик изготовления и анализа хромосомных препаратов. Только с появлением методик давленых, а позже и воздушно-высушенных препаратов, которые значительно превосходят по надежности и информативности метод реконструкции кариотипа по срезам, хромосомные числа у гидр стали определять более или менее точно. Стало ясно, что все изученные ранее гидры характеризуются многохромосомными и довольно консервативными по числу хромосом кариотипами. Проанализировав все собранные результаты и отбросив те, достоверность которых вызывала сомнения, мы пришли к выводу, что кариотипы H.viridissima и H.robusta включают, по всей вероятности, 30 хромосом. Тем же числом хромосом обладают, видимо, H.vulgaris и H.oligactis. Однако в отношении последних двух видов существуют некоторые разногласия между нашими результатами и данными других исследователей (одни авторы установили, что диплоидный набор у этих видов состоит из 30 хромосом, а другие — из 32). Это можно объяснить тем, что в обоих случаях под одним названием, видимо, скрывались разные виды, на что и указывают различия в числе хромосом. Хочется верить, что мы на правильном пути и, возможно, “нащупали” хороший признак для дифференциации близких видов у гидр. Но основные надежды мы возлагаем все же не на хромосомные числа, а на b- и g-кариологию. По известной классификации выдающегося цитогенетика М.Дж.Д.Уайта [9], хромосомный анализ на более высоких уровнях предусматривает изучение не только числа хромосом (a-уровень), но и детальных характеристик кариотипов. На b-уровне изучается морфология отдельных хромосом, т.е. локализация центромер и относительная длина хромосомных плеч; на g-уровне — индивидуальность рисунка дифференциального окрашивания хромосом. В настоящее время известно несколько типов дифференциального окрашивания хромосом по длине, однако не все они в равной мере воспроизводимы у разных организмов. Для гидр оказались перспективными методы, выявляющие структурный гетерохроматин и ядрышкообразующие районы хромосом. Была надежда, что удастся обнаружить межвидовые различия по количеству и локализации в хромосомах ядрышкообразующих районов (ЯОР) и сайтов С-гетерохроматина. Если бы мы добились успеха в использовании этих методов на нашем объекте, появились бы столь необходимые нам дополнительные маркеры для более точной идентификации гомологичных хромосом в каждом конкретном кариотипе и гомеологичных (соответствующих) хромосом при сравнительно-кариологическом анализе видов. В мечтах нам открывались замечательные возможности решения проблем систематики, эволюции, филогении гидр и, наконец, в перспективе — эволюции самих кариотипов. Мечты мечтами, но начинать надо было фактически с нуля. Знакомясь с работами предшественников, мы убедились, что надежной методики приготовления хромосомных препаратов из гидр нет, — качество метафазных пластинок, представленных на иллюстрациях, было из рук вон плохим. Начинать пришлось с разработки адекватной методики. Основная трудность заключалась в том, что у гидр для анализа хромосом, по общему мнению, наиболее пригодны интерстициальные клетки (т.е. клетки-основатели, способные развиваться в нервные, половые и стрекательные). Эти клетки не имеют в теле полипа конкретной локализации, поэтому найти их в достаточном количестве и, главное, в нужной фазе деления нам долго не удавалось. Для преодоления этой трудности мы испробовали разные ухищрения. Пытаясь принудить гидру к митозам, мы то кормили ее как на убой, то, наоборот, лишали пищи; без конца меняли режим кормления и температуру содержания. Разрезали ее на части, и тогда, в неистребимой жажде возрождения, она приступала к регенерации. Добавляли в солонку с гидрой фитогемагглютинин, широко применяемый для стимуляции клеточных делений у позвоночных животных, особенно млекопитающих. Чтобы увеличить выход метафазных пластинок, мы бесконечно экспериментировали с разными типами и концентрациями цитостатиков. Добиваясь хорошего разброса хромосом в метафазных пластинках, использовали разные гипотонические растворы и долго не могли решить, какие препараты нам нравятся больше — “давленые” или “воздушно-высушенные”. Наконец, мы добились некоторых успехов и даже опубликовали первые результаты. Так как жертвой наших жестоких экспериментов была H.vulgaris (та самая, которую считали H.attenuata), именно ей мы посвятили нашу первую статью о кариотипе гидр [10]. Это был первый вид гидр, у которого удалось выяснить локализацию ядрышкообразующего района в кариотипе. Мы отдавали себе отчет, что это — пока еще очень скромные результаты, однако бесконечно гордились, когда известный специалист по гидрам профессор Пьер Тардент, получив в подарок фотографию, высоко отозвался о качестве метафазной пластинки. Описанные эксперименты с гидрой принесли нам еще один чрезвычайно важный успех. Мы “нащупали” условия, при которых наша “экспериментальная” гидра переходила к половому размножению. Появилась возможность изучать хромосомы в мейозе. Теперь сперматогониальные деления обеспечивали нас обильным материалом для анализа. Наконец, можно было приступать к широкомасштабным исследованиям по кариосистематике гидр. Сейчас, когда мы пишем этот очерк, в разгаре полевой сезон. Мы собираем гидр и заготавливаем сотни воздушно-высушенных хромосомных препаратов, предвкушая наступление зимы, когда сможем их красить и изучать. Иногда нам не удается сдержать любопытства, и тогда в нашей лабораторной комнате горят лампочки термостатов (и — глаза исследователей), готовятся “хитрые” реактивы и в маленьких солонках ждут своей участи гидры, чьи кариотипы нам так не терпится увидеть. Таксономические исследования мы решили начать с гидр С.-Петербурга, Ленинградской обл., а затем северо- и юго-запада России и соседних территорий. Совершенно ясно, чем это объясняется: прежде всего доступностью материала и еще тем, что денег для полевых работ едва хватает на пригородные электрички... Правда, сейчас самый юный из авторов этой статьи путешествует по юго-западным озерам России и Белоруссии и, изучая гидр, подобно Трамбле, тратит собственные деньги. |
Есть ли у нас по прошествии года исследований какие-нибудь результаты? Да, есть. Вместо отмеченных в литературе для С.-Петербурга и Ленинградской обл. шести видов гидр мы обнаружили только три: H.oligactis, H.circumcincta и H.oxycnida. Иными словами, работая с живым материалом и используя надежные видовые критерии, мы вынуждены сокращать число реальных (валидных) видов гидр. Конечно же, мы были бы рады обнаружить в исследуемых регионах ранее не отмеченные виды. В дальнейшем нам бы хотелось получить ответ на вопрос о числе родов гидр, а также провести ревизию всего отряда Hydrida, в который многие исследователи до сих пор включают представителей семейства Protohydridae c одним или несколькими родами (такими как солоноватоводные Protohydra и даже морские Boreohydra). Кроме того, неплохо было бы выяснить, в каком родстве гидры находятся с представителями семейства Moerisiidae. Это — наши будущие планы, но уже сейчас мы готовы доказать ряд существенных различий между гидрами и протогидрами, последних мы решительно вынесли в другой отряд. Гидра населяет пресные водоемы самого разного типа: озера, реки и даже лужи, где есть растительность и достаточно чистая вода. Роль гидр в энергетическом балансе водоема изучена еще недостаточно, хотя есть публикации, в которых обсуждаются вопросы численности гидр, их пищевого рациона и механизмов питания [11]. По нашим данным, полученным при изучении популяции H.oxycnida в р.Оредеж, оказалось, что в 0.5 м3 воды может обитать до 2 тыс. особей, при этом потребляющих более 4500 мг ветвистоусых рачков в сутки. Это позволяет думать, что при массовом развитии гидры существенно влияют на продукцию зоопланктона в водоеме. Hо и этот вопрос требует более детального исследования. Что же получается?! Гидра, с которой начинается постижение животного мира в раннем школьном возрасте, гидра — модельный объект, на клетках и тканях которого производятся лабораторные эксперименты, а их результаты экстраполируются на решение общебиологических закономерностей, и при этом она остается практически неизученной в таксономическом и фаунистическом плане... Hеизвестно, сколько и каких видов гидр обитает в водоемах России и соседних регионов, какие особенности морфологии, микроанатомии, жизненных циклов и экологии следует рассматривать как основополагающие в таксономических построениях, какое место занимает гидра на эволюционном древе типа книдарий и какие из других таксонов следует ставить рядом с ней на этом древе, а какие, поставленные по соседству ранее, лучше было бы закрепить на других ветвях. Короче говоря, многое еще не известно про эту, такую знакомую гидру. Безусловно, в наши планы входит сбор и изучение фауны гидр всей России. Мы просим коллег из разных регионов собирать нам живых гидр. Hо вопрос с их пересылкой в Зоологический институт тоже решается с трудом. * * * Вольная форма изложения этого очерка располагает к ...анекдоту. Как
известно, баронесса фон Мекк существенно поддерживала материально творчество
П.И.Чайковского... “Что такое фон Мекк в жизни Чайковского?” — спрашивает
непосвященный работник Академии наук. “То же, что Фонд РФФИ в нашей
с тобой жизни”, — отвечает осведомленный коллега. Бог даст, так и будет....
Работа выполнена при поддержке РФФИ (грант 97-04-50100).
Литература 1 T r e m b l e y A. Memoires, pour servir a L’histoire d’un genre de polypes d’eau douce, a bras en forme de cornes. Leide, 1744; Т р а м б л е А. Мемуары к истории одного рода пресноводных полипов с руками в форме рогов / Пер. И.И.Канаева. М.; Л., 1937. 2 К а н а е в И.И. Гидра: Очерки по биологии пресноводных полипов. М.; Л., 1952. 3 H а у м о в Д.В. Гидроиды и гидромедузы
морских, солоноватоводных и пресноводных бассейнов СССР
4 L i n n a e u s K. Systema naturae. 1758. P.799—821. 5 P a l l a s P.S. Elenchus zoophytorum. Francofurti ad Moenum, 1766. 6 The biology of Hydra and of some other coelenterates / Eds H.M.Lenhoff, W.F.Loomis. Florida, 1961; From Tremley’s polyps to new directions in research on Hydra // Proc. Symposium Honoring Abraham Trembley (1710—1784) / Eds H.M.Lenhoff, P.Tardent. Univers. Californ. Irvine; Univers. Zurich; Soc. Phys. Hist. nat. Geneve. Florida, 1985; T a r d e n t P. Hydra. Zurich, 1988; H o l s t e i n T.W. Cnidaria: Hydrozoa. Susswasserfauna von Mitteleuropa. Jena, 1995. 7 B r i e n P. L’embriogenese et la senescence de l’hydre d’eau douce //Mem. de l’Acad. Roy. Belg. Sci. 1965. T.36. P.1—113. 8 B r a u e r A. U``ber die Entwicklung von Hydra // Zeitschr. Wiss. Zool. 1891. Bd.52. S.169—216. 9 W h i t e M.J.D. Modes of speciation. San Francisco, 1978. 10 О в а н е с я н И.Г., К у з н е ц о в а В.Г. // Книдарии. Современное состояние и перспективы исследований. Вып.II / Под ред. С.Д.Степаньянц // Тр. Зоол. ин-та. 1995. Т.261. С.96—102. 11 C u k e r B.E., M o z l e y S.C. // Limnol. Oceanogr. 1981. V.26. № 4. Р.697—708; Б а б и ц к и й В.А. // Зоол. журн. 1995. Т.74. Вып.4. С.22—33. |