№ 8-9, 2001 г.
№ 8-9, 2001 г.
№ 8-9, 2001 г.
№ 8-9, 2001 г.
© М.Д.ГолубовскийНеканонические наследственные изменения
М.Д. Голубовский
Михаил Давидович Голубовский, доктор биологических наук, ведущий научный сотрудник
Санкт-Петербургского филиала Института истории естествознания и техники РАН.Генетика как наука оформилась 100 лет назад, после вторичного открытия законов Менделя. Ее бурное развитие ознаменовалось в последние годы расшифровкой нуклеотидного состава ДНК генома многих десятков видов. Возникли новые ветви знаний — геномика, молекулярная палеогенетика. В начале 2001 г. в рамках дорогостоящей 10-летней международной программы объявлено о принципиальной расшифровке генома человека. Эти достижения, пожалуй, можно сравнить с выходом человека в космос и высадкой на Луну.
Генетическая инженерия и биотехнология сильно изменили облик науки. Вот любопытный эпизод, уже попавший в новейшую сводку: “После 1998 года началась беспрецедентная гонка между 1100 учеными мирового сообщества проекта “Геном человека” и частной акционерной фирмой “Celera Genomics”” [1]. Фирма надеялась первой прийти к финишу и извлекать выгоду от патентования фрагментов ДНК человека. Но пока победил принцип: “Что создано природой и Богом, не может патентоваться человеком”.
Мог ли представить такую фантасмагорическую картину Грегор Мендель, неспешно проводя год за годом свои опыты в тиши монастырского садика? В какой степени она трансформирует естественное саморазвитие науки? Действительно ли тотальный анализ ДНК геномов снимает все покровы? Упования, что Буратино уже нашел заветный золотой ключик от потайной двери, столкнулись с непредвиденной реальностью и парадоксами. У человека лишь 3% ДНК генома кодируют белки, и, возможно, еще 20—25% участвуют в регуляции действия генов. Какова же функция, и есть ли она у остальной части ДНК? Гены в геноме порой сравнивают с небольшими островами в море неактивных и, возможно, “мусорных” последовательностей. ДНК-гонка порой напоминает присказку: “принеси то, не знаю что”.
Возражения скептиков отнюдь не сняты. Ведь при тотальном секвенировании номинация (применю модный термин) определенного отрезка ДНК в “ранг гена” проводится лишь на основе сугубо формальных критериев (знаков генетической пунктуации, необходимых для транскрипции). Роль, время и место действия большинства “генов-номинантов” пока совершенно неясны.
Но есть и другая проблема. Под геномом надо понимать всю наследственную систему, включая не только структуру определенного набора ДНК элементов, но и характер связей между ними, который определяет ход онтогенеза в конкретных условиях среды. Налицо системная триада: элементы, связи между ними и свойства целостности. Отсюда следует важный вывод: знание структуры генов на уровне ДНК — необходимо, но вовсе недостаточно для описания генома. Мы лишь на пороге постижения динамического способа организации и неканонических форм наследования [2, 3].
Неожиданно в конце ХХ в. вопрос о том, каковы границы и спектр наследственной изменчивости, вышел за рамки чисто академических дискуссий. Сначала в Англии, а потом и в Германии пришлось забивать скот из-за нейродегенеративной аномалии, способной передаваться людям с мясом больных животных. Инфекционным агентом оказались не ДНК или РНК, а белки, названные прионами (от англ. prions — protein infectious particles — белковые инфекционные частицы).
Впервые с их необычным проявлением исследователи столкнулись еще в 60-е годы. Но тогда этот феномен пытались истолковать в рамках классических представлений, полагая, что это “медленные вирусные инфекции” животных или особый тип супрессорных мутаций у дрожжей. Теперь выясняется, “феномен прионов не является экзотикой, характерной для млекопитающих, а скорее — частным случаем общебиологического механизма” динамического наследования [4]. Вероятно, центральную догму молекулярной генетики придется дополнить с учетом возможности внутри- и межвидовой передачи по типу инфекций [5].
В начале 80-х годов классик молекулярной биологии и генетики Р.Б.Хесин выделил три формы неканонической наследственной изменчивости: неслучайные упорядоченные изменения в локусах и участках хромосом, состоящих из повторов ДНК; изменение и наследование свойств цитоплазмы; эпигенетическое наследование локальных и общих изменений упаковки хроматина. Затем добавились мобильные гены, поведение которых привело к проблеме непостоянства геномов [6].
Цель этой статьи — показать, что разные формы неменделевского наследования не исключение, а следствие более общих представлений об организации генома. Наследственные изменения отнюдь не сводятся только к мутациям [7].
Андре Львов и роль его открытия
По удивительному совпадению в один и тот же 1953 г. появились две статьи, определившие лицо современной генетики: открытие двойной спирали ДНК Дж.Уотсоном и Ф.Криком и концепция профага и лизогении бактерий А.Львова (1902—1994), которая, на мой взгляд, имеет сейчас для биологии, медицины и генетики не меньшее значение, чем двойная спираль ДНК.
Львов установил, что фаг может встраиваться в хромосому бактерии и передаваться в ряду многих поколений как обычный бактериальный ген. В таком состоянии у фага работает лишь ген-репрессор, который блокирует работу всех остальных его локусов. Бактерия, включившая фаг в свой геном, называется лизогенной, а встроенный фаг — профагом. Такая лизогенная бактерия защищена от заражения другими фагами. Под действием ультрафиолета или изменений внутренней среды клетки репрессор инактивируется, блокада снимается, и фаг размножается, вызывая гибель клетки. Теперь даже трудно представить, сколь революционным было это открытие.
Андре Львов — выходец из России, его родители эмигрировали во Францию в конце XIX в. Образ матери ученого Марии Симинович навсегда запечатлен на полотне художника В.Серова “Девушка, освещенная солнцем” (1888). Мария Яковлевна Львова-Симинович дожила до 90 лет. За несколько недель до второй мировой войны она передала в Третьяковскую галерею письма и рисунки В.Серова [9]. Отец Львова был знаком с Мечниковым и водил сына к нему в Институт Пастера. Так через века и страны тянутся и переплетаются нити культуры. За свою долгую жизнь А.Львов работал последовательно как протозоолог, бактериолог, биохимик, генетик и, наконец, как вирусолог. В Институте Пастера он патронировал и Ж.Моно, и Ф.Жакобу, разделивших с мэтром Нобелевскую премию 1965 г. за открытие оперона.
Уже с 20-х годов были известны штаммы бактерий, которые якобы несут фаги в скрытом состоянии и время от времени вызывают лизис клеток. Однако открыватель бактериофагии Ф.Д'Эррель смотрел на фаг только как на летальный для клетки агент, не допуская мысли о его скрытом состоянии. Это мнение разделял сначала и классик молекулярной генетики М.Дельбрюк. Дело в том, что он и его коллеги в США работали с так называемыми Т-фагами, которые неспособны встраиваться в хромосому бактерий. В силу “демона авторитетов” лизогенией с 20-х годов скрупулезно не занимались. Пионер этих работ, блестящий микробиолог из Института Пастера, Эжен Вольман был схвачен немцами как еврей во время оккупации Парижа и погиб.
После войны Львов возобновил в Пастеровском институте исследования скрытого фагоносительства. В 1953 г. он создал стройную концепцию профага, сразу же осознав ее значение для вирусной теории рака и ряда вирусных патологий у человека. Его ясная схема феномена лизогении до сих пор приводится во всех сводках по молекулярной генетике.
В 1958 г. Ф.Жакоб и Элиас Вольман (сын Эжена Вольмана) ввели термин “эписома” для элементов, способных существовать либо в свободном, либо интегрированном в геном хозяина состоянии. К эписомам они отнесли умеренные фаги, половой фактор бактерий, факторы колициногенности, с помощью которых одни штаммы бактерий убивают другие бактерии. В замечательной книге “Пол и генетика бактерий”, написанной в 1961 г. (и уже на следующий год вышедшей стараниями известного генетика С.И.Алиханяна в русском переводе), авторы предвидели существование эписомоподобных элементов и у высших организмов, прозорливо указывая на “контролирующие элементы”, открытые Б.Мак-Клинток в начале 50-х годов (Нобелевская премия по физиологии и медицине 1983 г.). Однако в то время они не предполагали, сколь глубока эта аналогия. После открытия в начале 70-х годов инсерционных мутаций, вызванных включением вирусной ДНК в клеточный геном бактерий, появилась возможность выстроить эволюционный ряд двусторонних переходов: инсерционные сегменты“транспозоны“плазмиды“фаги.
Сходные ряды перевоплощений найдены и у эвкариот. У дрозофилы мобильные элементы семейства gypsy (“цыган”) могут существовать в виде встроенных в хромосому копий; находиться в виде их полных или уменьшенных кольцевых или линейных плазмид в цитоплазме; наконец, в случае отдельных “разрешающих” мутаций в геноме хозяина способны одеться оболочкой, стать настоящими инфекционными ретровирусами и заражать посторонних хозяев через корм. Сходство Р-транспозонов у дрозофилы и эндогенного ретровируса ВИЧ у человека (табл.) позволяет предсказывать возможные эволюционно-генетические события в популяциях человека, судьбу неизбежных сейчас и в будущем его контактов с чужими геномами.
Принцип факультативности и обобщенная концепция генома
Многие факты изменчивости, связанной с мобильными элементами, не укладываются в представления о мутациях как о локализованных изменениях структуры, числа или расположения генных локусов. Дабы совместить данные классической и “подвижной” генетики, я в 1985 г. предложил естественную классификацию элементов генома, включающую две подсистемы: облигатные (гены и их регуляторные области в хромосомах) и факультативные элементы (ДНК- и РНК-носители, число и топография которых варьирует в разных клетках или организмах одного вида) [8].
Из этой классификации вытекают важные следствия, позволяющие осмыслить или сформулировать многие необычные факты из области наследственной изменчивости. Назовем некоторые из них:
Мак-Клинток впервые сделала заключение, что активация факультативных элементов и последующая структурная реорганизация генома могут быть следствием адаптивного ответа клетки на стресс [9]. Наследственная система, активируя факультативные элементы, осуществляет генетический поиск, переходя к новому адаптивному уровню функционирования. Так, многолетние исследования Л.З.Кайданова показали, что после длительного инбридинга в линиях дрозофил вдруг за одно-два поколения происходят множественные кооперативные перемещения подвижных генов и сайт-специфичные перестройки хромосом; при этом одновременно резко повышается выживаемость.
- универсальность факультативности. Нет видовых геномов, которые состоят лишь из облигатных элементов, как нет живых организмов, состоящих лишь из скелетного остова;
- генетическая неидентичность дочерних клеток. В силу случайности они различаются по числу и составу цитоплазматических факультативных элементов. Соотношение фракций облигатных и факультативных элементов ДНК – относительно устойчивый видовой признак. Имея сходное число генных локусов, близкие виды могут отличаться по количеству ДНК в 2—5 и более раз, наращивая блоки повторов и меняя их геномную топографию. Между облигатной и факультативной частями генома непрерывно наблюдаются разные переходы. Самые очевидные примеры — мутации генов за счет внедрения (инсерций) мобильных элементов или умножение числа (амплификация) сегментов хромосом и переход их в разные внутри- и внехромосомные состояния;
- характерный тип наследственной изменчивости для каждой из двух подсистем генома. Моргановские мутации легко соотносятся с облигатным компонентом. Разнообразные наследственные изменения в числе и топографии факультативных элементов я предложил называть “вариациями” (как в музыке — вариации на заданную тему). Мутации, согласно классическим представлениям, происходят, как правило, случайно, с низкой частотой у отдельных особей. Характер вариаций совсем иной — здесь возможны массовые, упорядоченные изменения под действием самых разных, в том числе слабых немутагенных факторов (температуры, пищевого режима и т.д.);
- двухэтапный характер большинства природных наследственных изменений. Сначала активизируются факультативные элементы как наиболее чувствительные к изменениям среды. Затем начинают опосредованно поражаться и генные локусы. К такому выводу мы пришли в ходе многолетних наблюдений за вспышками мутаций в природе. Большинство их оказалось нестабильными и были вызваны инсерциями мобильных элементов, которые таинственным образом время от времени активируются в природе. У дрозофилы около 70% мутаций, возникших в природе или лаборатории спонтанно, связано с перемещениями мобильных элементов.
Обобщенное представление о геноме как ансамбле облигатных и факультативных элементов расширяет и понятие “горизонтальный перенос”, включающий не только интеграцию чужеродных генов в хромосомы ядра. О горизонтальном переносе можно говорить уже и в случаях создания устойчивой ассоциации двух генетических систем, в которой появляются новые признаки и свойства.
Функциональная факультативность генома
Наследственные изменения возникают в результате ошибок процессов, оперирующих с наследственным материалом любых живых организмов, — репликации, транскрипции, трансляции, а также репарации и рекомбинации.
Факультативность репликации означает возможность относительно автономной гипер- или гипорепликации отдельных участков ДНК, независимо от плановой закономерной репликации всей геномной ДНК в ходе деления клетки. Такими свойствами обладают участки хромосом с повторами, блоки гетерохроматина. При этом автономная репликация приводит к умножению числа отдельных сегментов и имеет, как правило, адаптивный характер.
Факультативность транскрипции состоит в возможности появления разных мРНК с одной и той же матрицы за счет присутствия в данном локусе более одного промотора и альтернативного сплайсинга. Эта ситуация нормальна для множества генов.
Неоднозначность (по терминологии С.Г.Инге-Вечтомова) трансляции проявляется в разных вариантах опознания одного и того же кодона, например стоп-кодона или кодона для включения определенной аминокислоты в синтезируемый белок. Такая трансляция зависит от физиологических условий в клетке и от генотипа.
Согласно теории мутационного процесса М.Е.Лобашева, возникновение мутации связано со способностью клетки и ее наследственных структур к репарации повреждений. Отсюда следует, что появлению мутации предшествует состояние, когда повреждение либо полностью обратимо, либо может реализоваться в виде мутации, понимаемой как “нетождественная репарация”. К началу 70-х годов выяснилось, что стабильность ДНК в клетке не имманентное свойство самих молекул ДНК, — оно поддерживается особой ферментативной системой.
С середины 70-х годов стала проясняться эволюционная роль “ошибок рекомбинации” как индуктора наследственных изменений, причем гораздо более мощного, чем ошибки репликации ДНК.
На молекулярном уровне различают три варианта рекомбинации: общую, сайт-специфичную и репликативную. Для первой, общей, регулярной рекомбинации (кроссинговер) репарация включают разрывы в цепи ДНК, их сшивку и восстановление. Для нее необходимы длинные районы гомологии ДНК. Сайт-специфичная рекомбинация довольствуется короткими, в несколько оснований, участками гомологии, какие, к примеру, имеют ДНК фага l и хромосома бактерии. Аналогично происходит включение в геном мобильных элементов и соматическая локальная рекомбинация в онтогенезе между иммуноглобулиновыми генами, создающая их поразительное разнообразие.
Ошибки общей рекомбинации можно рассматривать как закономерные следствия линейно протяженной структуры генов. Возникает дилемма, о которой писал Хесин: можно считать, что митотические рекомбинации — особый тип мутагенеза или, напротив, некоторые виды мутаций (хромосомные аберрации) — результат “ошибок” митотических рекомбинаций.
Если перемещения мобильных элементов или рекомбинация участков запрограммированы в онтогенезе, классифицировать такие наследственные изменения трудно. Трансформацию пола у дрожжей долгое время считали мутационным событием, но оказалось, что на определенной стадии развития аскоспор она происходит с высокой вероятностью в результате сайт-специфичной рекомбинации.
Вариации генома в ответ на вызов среды
В теории эволюции и в генетике всегда обсуждался вопрос о связи наследственных изменений с направлением отбора. Согласно дарвиновским и постдарвиновским представлениям, наследственные изменения происходят в разных направлениях и лишь затем подхватываются отбором. Особенно наглядным и убедительным оказался метод реплик, изобретенный в начале 50-х годов супругами Ледерберг. С помощью бархатной материи они получали точные копии — отпечатки — опытного посева бактерий на чашке Петри. Затем на одной из чашек вели отбор на устойчивость к фагу и сопоставляли топографию точек появления устойчивых бактерий на чашке с фагом и в контроле. Расположение устойчивых к фагу колоний было одинаковым в двух чашках-репликах. Такой же результат получили и при анализе положительных мутаций у бактерий, дефектных по какому-либо метаболиту.
Открытия в области подвижной генетики показали, что клетка как целостная система в ходе отбора может адаптивно перестраивать свой геном. Она способна ответить на вызов среды активным генетическим поиском, а не пассивно ждать случайного возникновения мутации, позволяющей выжить. А в опытах супругов Ледерберг у клеток не было выбора: либо смерть, либо адаптивная мутация.
В тех же случаях, когда фактор отбора не летален, возможны постепенные перестройки генома, прямо или косвенно связанные с условиями отбора. Это выяснилось с открытием в конце 70-х годов постепенного умножения числа локусов, в которых расположены гены устойчивости к селективному агенту, блокирующему деление клеток. Известно, что метотрексат — ингибитор клеточного деления — широко применяется в медицине для остановки роста злокачественных клеток. Этот клеточный яд инактивирует фермент дигидрофолатредуктазу (ДГФР), работу которого контролирует определенный ген.
Рассмотрим, как в ходе подобного отбора менялся геном у одноклеточного паразитического жгутиконосца лейшмании Leishmania tropica, вызывающего кожные язвы и передающегося человеку москитами от грызунов. До начала опытов ген устойчивости был в геноме лейшмании в одной дозе. В результате отбора число генов устойчивости увеличилось (амплифицировалось). Что же наблюдали при разных вариантах отбора?
Устойчивость клеток лейшмании к яду-цитостатику (метотрексат) возрастала ступенчато, и пропорционально увеличивалась доля амплифицированных сегментов с геном устойчивости. Умножался не только селектируемый ген, но и большие прилежащие к нему участки ДНК, названные ампликонами. Когда устойчивость к яду у лейшмании повысилась в 1000 раз, амплифицированные внехромосомные сегменты составили до 10% ДНК в клетке! Можно сказать, что из одного облигатного гена образовался пул факультативных элементов. Произошла адаптивная перестройка генома в ходе отбора.
Если отбор продолжался достаточно долго, часть ампликонов встраивалась в исходную хромосому, и после прекращения отбора устойчиво сохранялась повышенная устойчивость.
С удалением из среды селективного агента число ампликонов с геном устойчивости постепенно снижалось в ряду поколений и одновременно падала устойчивость. Тем самым был смоделирован феномен длительных модификаций, когда массовые изменения, вызванные средой, наследуются, но постепенно угасают в ряду поколений.
При повторном отборе часть сохранившихся в цитоплазме ампликонов обеспечивала быструю их автономную репликацию, и устойчивость возникала гораздо быстрее, чем в начале опытов. Иными словами, формировалась своеобразная клеточная ампликонная память о прошедшем отборе на основе сохранившихся ампликонов.
Если сопоставить метод реплик и ход отбора на устойчивость в случае амплификации, то оказывается, что именно контакт с селективным фактором вызвал преобразование генома, характер которого коррелировал с интенсивностью и направлением отбора.
Дискуссия об адаптивных мутациях
В 1988 г. в журнале “Nature” появилась статья Дж.Кэйрнса с соавторами о возникновении у бактерии E.coli отборзависимых “направленных мутаций”. Брали бактерии, несущие мутации в гене lacZ лактозного оперона, неспособные расщеплять дисахарид лактозу. Но эти мутанты могли делиться на среде с глюкозой, откуда их через один—два дня роста переносили на селективную среду с лактозой. Отобрав lac+ реверсов, которые, как и ожидалось, возникли еще в ходе “глюкозных” делений, нерастущие клетки оставляли в условиях углеводного голодания. Сначала мутанты отмирали. Но спустя неделю и более наблюдался новый рост за счет вспышки реверсий именно в гене lacZ. Как будто клетки в условиях жесткого стресса, не делясь (!), вели генетический поиск и адаптивно меняли свой геном [10].
В последующих работах Б.Холла использовались бактерии, мутантные по гену утилизации триптофана (trp). Их помещали на среду, лишенную триптофана, и оценивали частоту реверсий к норме, которая повышалась именно при триптофановом голодании. Но причиной этого феномена были не сами условия голодания, ибо на среде с голоданием по цистеину частота реверсий к trp+ не отличалась от нормы.
В следующей серии опытов Холл взял уже двойных недостаточных по триптофану мутантов, несущих одновременно мутации в генах trpA и trpВ, и вновь поместил бактерии на среду, лишенную триптофана. Выжить могли только особи, у которых реверсии возникали одновременно в двух триптофановых генах. Частота появления таких особей была в 100 млн раз выше, чем ожидалось при простом вероятностном совпадении мутаций в двух генах. Холл предпочел называть этот феномен “адаптивные мутации” и впоследствии показал, что они возникают и у дрожжей, т.е. у эвкариот [11].
Публикации Кэйрнса и Холла немедленно вызвали бурную дискуссию. Итогом ее первого раунда стало выступление одного из ведущих исследователей в области подвижной генетики Дж.Шапиро. Он кратко обсудил две основные идеи. Во-первых, клетка содержит биохимические комплексы, или системы “естественной генетической инженерии”, которые способны реконструировать геном. Активность этих комплексов, как и любая клеточная функция, может резко меняться в зависимости от физиологии клетки. Во-вторых, частота возникновения наследственных изменений всегда оценивается не для одной клетки, а для клеточной популяции, в которой клетки могут обмениваться между собой наследственной информацией. Кроме того, межклеточный горизонтальный перенос с помощью вирусов или передачи сегментов ДНК усиливается в стрессовых условиях. Как считает Шапиро, эти два механизма объясняют феномен адаптивных мутаций и возвращают его в русло обычной молекулярной генетики [11]. Каковы же, на его взгляд, итоги дискуссии? “Мы нашли там генетического инженера с впечатляющим набором замысловатых молекулярных инструментов для реорганизации ДНК-молекулы” [12].
За последние десятилетия на уровне клетки открыта такая непредвиденная сфера сложности и координации, которая более совместима с компьютерной технологией, нежели с механизированным подходом, доминировавшим во время создания неодарвинистского современного синтеза. Вслед за Шапиро, можно назвать по крайней мере четыре группы открытий, изменивших понимание клеточных биологических процессов.
Организация генома. У эвкариот генетические локусы устроены по модульному принципу, представляя собой конструкции из регуляторных и кодирующих модулей, общих для всего генома. Это обеспечивает быструю сборку новых конструкций и регуляцию генных ансамблей. Локусы организованы в иерархические сети, во главе с главным геном-переключателем (как в случае регуляции пола или развития глаза). Причем многие из соподчиненных генов интегрированы в разные сети: они функционируют в разные периоды развития и влияют на множество признаков фенотипа.В условиях вызова среды клетка действует целенаправленно, подобно компьютеру, когда при его запуске шаг за шагом проверяется нормальная работа основных программ, и в случае неисправности работа компьютера останавливается. В целом становится очевидной, уже на уровне клетки, правота нетрадиционного французского зоолога-эволюциониста Поля Грассэ: “Жить — значит реагировать, а отнюдь не быть жертвой”.Репаративные возможности клетки. Клетки вовсе не пассивные жертвы случайных физико-химических воздействий, поскольку в них имеется система репараций на уровне репликации, транскрипции и трансляции.
Мобильные генетические элементы и природная генетическая инженерия. Работа иммунной системы построена на непрерывном конструировании новых вариантов молекул иммуноглобулинов на основе действия природных биотехнологических систем (ферменты: нуклеазы, лигазы, обратные транскриптазы, полимеразы и т.д.). Эти же системы используют мобильные элементы для создания новых наследуемых структур. При этом генетические изменения могут быть массовыми и упорядоченными. Реорганизация генома — один из основных биологических процессов. Природные генноинженерные системы регулируются системами с обратной связью. До поры до времени они пребывают в неактивном состоянии, но в ключевые периоды или во время стресса приводятся в действие.
Клеточный информационный процессинг. Возможно, одно из самых важных открытий в области биологии клетки состоит в том, что клетка непрерывно собирает и анализирует информацию о своем внутреннем состоянии и внешней среде, принимая решение о росте, движении и дифференциации. Особенно показательны механизмы контроля клеточного деления, лежащие в основе роста и развития. Процесс митоза универсален у высших организмов и включает три последовательных этапа: подготовка к делению, репликация хромосом и завершение деления клетки. Анализ генного контроля этих фаз привел к открытию особых точек, в которых клетка проверяет, произошла ли репарация нарушений в структуре ДНК на предыдущем этапе или нет. Если ошибки не будут исправлены, последующий этап не начнется. Когда же ликвидировать повреждения нельзя, запускается генетически запрограммированная система клеточной смерти, или апоптоза.
Пути возникновения естественных наследственных изменений в системе среда—факультативные элементы—облигатные элементы. Факультативные элементы первыми воспринимают немутагенные факторы среды, а возникающие затем вариации вызывают мутации. На поведение факультативных элементов влияют и облигатные элементы.
Неканонические наследственные изменения, возникающие под влиянием отбора к цитостатикам и приводящие к амплификации генов.
Приобретенные признаки наследуются
“История биологии не знает более выразительного примера многовекового обсуждения проблемы, чем дискуссия о наследовании или о ненаследовании приобретенных признаков”, - эти слова стоят в начале книги известного цитолога и историка биологии Л.Я.Бляхера [12]. В истории, пожалуй, можно вспомнить аналогичную ситуацию с попытками превращения химических элементов. Алхимики верили в эту возможность, но в химии утвердился постулат о неизменности химических элементов. Однако ныне в ядерной физике и химии исследования по превращению элементов и анализ их эволюции - дело обычное. Кто же оказался прав в многовековом споре? Можно сказать, что на уровне химических молекулярных взаимодействий не происходит превращения элементов, а на ядерном уровне оно - правило.
Напрашивается сходная аналогия и с вопросом о наследовании признаков, которые появились в ходе онтогенеза. Если вновь возникающие наследственные изменения сводить лишь к мутациям генов и хромосом, тогда вопрос можно считать закрытым. Но если исходить из обобщенной концепции генома, включая представление о динамической наследственности [2, 3], проблема нуждается в пересмотре. Помимо мутационной существует вариационная и эпигенетическая формы наследственной изменчивости, связанные не с изменениями в тексте ДНК, а в состоянии гена. Такие эффекты обратимы и наследуемы.
Интересно, что вышедший в конце 1991 г. Международный ежегодник по генетике открывается статьей О.Ландмана “Наследование приобретенных признаков” [13]. Автор суммирует уже давно полученные в генетике факты, показывая, что “наследование приобретенных признаков вполне совместимо с современной концепцией молекулярной генетики”. Ландман детально рассматривает около десяти экспериментальных систем, в которых установлено наследование приобретенных признаков. Четыре разных механизма способны привести к нему: изменение структур клеточной оболочки, или кортекса, изученное Т. Соннеборном у инфузорий; ДНК-модификации, т.е. клонально передаваемые изменения в характере локального метилирования ДНК (сюда входит феномен импринтинга); эпигенетические изменения без каких-либо модификаций ДНК; индуцированная утрата либо приобретение факультативных элементов.
Статья Ландмана делает нас как бы свидетелями критического периода смены постулата в генетике, казавшегося неколебимым как скала. Автор спокойно, без ажиотажа и новых сногсшибательных фактов, объединяет старые и новые данные в систему, дает им ясное современное истолкование. Можно сформулировать общий принцип: наследование приобретенных признаков возможно в тех случаях, когда некий фенотипический признак зависит от числа или топографии факультативных элементов.
Приведу два поучительных примера на дрозофиле: первый связан с поведением вируса сигма, второй - мобильных элементов, ответственных за гибридную стерильность самок и сверхмутабильность.
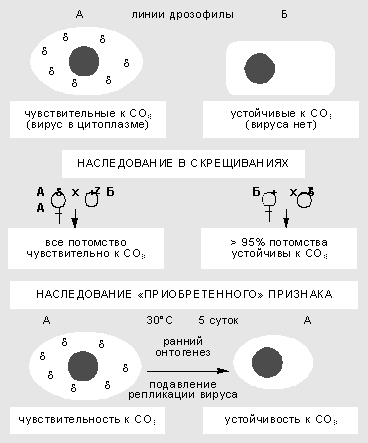
Изучение взаимодействия вируса сигма с геномом дрозофилы началось более 60 лет назад. Сначала в 1937 г. французский генетик Ф.Леритье обнаружил резкие наследственные отличия у разных линий мух по степени чувствительности к углекислому газу (СО2). Признак наследовался причудливым образом: через цитоплазму, но не только по материнской линии, а иногда и через самцов. Чувствительность можно было передать и путем инъекции гемолимфы, причем разным видам дрозофил. В этих случаях признак передавался не стабильно, но в результате отбора наследование становилось устойчивым.
Неменделевское наследование признака у дрозофилы, который зависит от популяции факультативных элементов генома. Признак чувствительности к СО2 вызван присутствием в цитоплазме мухи рабдовируса сигма. В результате температурного шока на ранней стадии развития дрозофилы блокируется размножение вируса, и выросшие особи приобретают устойчивость к нему.Чувствительность к СО2 оказалась связана с устойчивым размножением в половых и соматических клетках РНК-содержащего пулевидного рабдовируса сигма, сходного по ряду свойств с вирусом бешенства у млекопитающих. Оогонии (клетки, из которых в ходе мейоза и созревания образуются яйцеклетки) у самок стабилизированной линии обычно содержат 10-40 вирусных частиц, а ооциты (зрелые яйцеклетки) - 1-10 млн. Вирус сигма - типичный факультативный элемент. Мутации в его геноме приводят к сложным формам поведения системы. Найдены случаи вирусоносительства, при которых дрозофилы остаются устойчивы к СО2, но вместе с тем иммунны к заражению другими штаммами вируса. Ситуация вполне сравнима с поведением системы фаг-бактерия, что сразу заметили Ф.Жакоб и Э.Вольман.Взаимоотношения генома дрозофилы и размножающегося в ее цитоплазме вируса подчиняются правилам внутриклеточной генетики. Воздействия в ходе онтогенеза могут вызвать сдвиг в числе и межклеточной топографии частиц и как результат - изменить степень чувствительности к углекислому газу. Так, повышенная температура блокирует репликацию вирусных частиц. Если самок и самцов в период гаметогенеза содержать несколько дней при температуре 30°С, потомство от таких мух будет свободно от вируса и устойчиво к СО2. Значит, приобретенный в ходе индивидуального развития признак наследуется в ряду поколений.
Ситуация с вирусом сигма не единична. Французские генетики изучали факторы стерильности самок, связанные с поведением мобильных элементов типа “I”. Наследование этого признака определяется сложными ядерно-цитоплазматическими взаимодействиями. Если в отцовских хромосомах локализованы активные I-элементы, то на фоне R-цитоплазмы они начинают активироваться, претерпевают множественные транспозиции и в результате вызывают резкие нарушения онтогенеза в потомстве самок с чувствительной цитоплазмой. Такие самки откладывают яйца, но часть эмбрионов гибнет на ранней стадии дробления - еще до образования бластомеры. Линии, выделенные из природных популяций, отличаются по силе действия I-факторов и степени реактивности (или чувствительности) цитоплазмы. Эти показатели можно изменить внешним влиянием. Возраст исходных родительских самок, а также воздействие в ранний период развития повышенной температуры отражаются не только на плодовитости выросших самок, но и на плодовитости их потомства. Вызванные условиями среды изменения реактивности цитоплазмы поддерживаются на протяжении многих клеточных поколений. “Самое замечательное, что эти изменения реактивности цитоплазмы под влиянием негенетических факторов наследуются: наблюдается наследование “благоприобретенных” признаков”, - отмечал Р.Б.Хесин [5].
Наследование через цитоплазму: от бабушек до внуков
В теории развития и феногенетике ХХ в. важное место занимают глубокие и совершенно оригинальные исследования эмбриолога П.Г.Светлова (1892-1972). Остановимся на разработанной им теории квантованности онтогенеза (наличии критических периодов в развитии, когда происходит детерминация морфогенетических процессов и одновременно повышается чувствительность клеток к повреждающим агентам) и на развитой в связи с этим идее, что изучение онтогенеза надо вести не с момента оплодотворения и образования зиготы, а еще с гаметогенеза, включающего оогенез у самок предшествующего поколения - проэмбрионального периода.
На основании этих постулатов Светлов провел в 60-е годы простые и ясные опыты на дрозофиле и мышах. Он убедительно показал, что возможно стойкое неменделевское наследование свойств цитоплазмы, а модификации в выраженности мутантных признаков, возникшие после кратковременного внешнего воздействия в критический период развития организма, тоже передаются в ряду поколений [14].
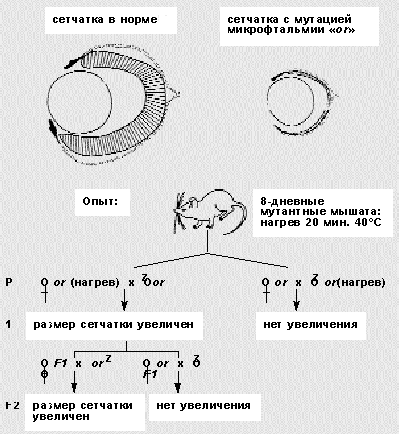
В одной из серий опытов он сравнивал степень проявления мутантного признака в потомстве двух линий мышей, гетерозиготных по рецессивной мутации микрофтальмии (уменьшенный размер сетчатки и глаз с момента рождения): нормальных по фенотипу гетерозигот, у которых мутантными были матери, и тех, у которых мутантны отцы. Потомство от мутантной бабушки отличалось более сильным проявлением признака. Светлов объяснял этот странный факт тем, что женские гаметы гетерозиготных самок находились еще в теле их мутантных матерей и испытывали с их стороны влияние, которое усилило мутации у внуков.
По существу Светлов установил явление, впоследствии получившее название “геномный импринтинг” - различие в выраженности гена в зависимости от того, пришел он к потомству от матери или от отца. Работы эти, увы, остались недооцененными.
Интересно, что еще в конце 80-х годов импринтинг, как остроумно заметил К.Сапиенца, исследователь этого феномена, было “принято считать генетическим курьезом, затрагивающим лишь очень немногие признаки. Меня неоднократно спрашивали, почему я попросту трачу свое время на столь незначительное явление” [15]. Большинство исследователей безоговорочно принимали одно из главных положений Менделя - “зачаток”, или ген, не может менять свои потенции в зависимости от пола, на чем основано повсеместно наблюдаемое расщепление 3:1. Но Сапиенца совершенно справедливо заметил, что при анализе менделевского расщепления обычно рассматривают только наличие или отсутствие признака, а если он количественный, то границу есть-нет устанавливают по принятому порогу. Если же выявить, какова степень проявления признака, обнаружится влияние геномного импринтинга.
Именно таков был подход Светлова, когда он тщательно изучал, как меняется выраженность признаков у потомства в зависимости от материнского генотипа. Как эмбриолог он видел общность наследственных и особых ненаследственных изменений - фенокопий (имитирующих мутации), если затрагивается один и тот же морфогенетический аппарат, ответственный за осуществление данного признака.
Впервые на разных видах животных (дрозофилах и мышах) Светлов показал возможность наследования через мейоз измененного характера проявления мутантного гена. Недаром эти работы Хесин в своей сводке назвал замечательными [16].
Кратковременное (20 мин) прогревание тела восьмидневного мышонка самки вызывало стойкие изменения ооцитов, ослаблявшие действие вредной мутации у внуков! “Передача улучшения развития глаз, наблюдаемая в опытах с нагреванием, может быть объяснена только передачей свойств, приобретенных ооцитами нагретых самок по наследству” [16]. Этот феномен Светлов связывал с особенностями формирования и строения яйцеклетки у животных, ибо “в ооците имеется как бы каркас, отражающий наиболее общие черты архитектоники строящегося организма”. Для профилактики нарушений развития у человека он обосновал необходимость изучения критических периодов гаметогенеза, в которых повышена чувствительность к повреждениям. Возможно, в патогенезе аномалий развития у человека этап формирования гамет даже более важен, чем эмбриогенез.
Схема опытов П.Г.Светлова, демонстрирующих передачу в ряду поколений мышей мутации - микрофтальмии. Однократное 20-минутное действие повышенной температуры на мутантных восьмидневных мышат влечет улучшение развития глаз у их потомков (F1 и F2). Этот признак наследуется только по материнской линии и связан с изменением в ооцитах.Сегодня этот вывод подтвержден молекулярно-генетическими исследованиями последнего десятилетия [17]. У дрозофилы установлены три системы материнских генов, которые формируют осевую и полярную гетерогенность цитоплазмы и градиенты распределения биологически активных генных продуктов. Задолго до начала оплодотворения происходит молекулярная детерминация (предопределение) плана строения и начальных этапов развития. В формировании ооцита большую роль играют генопродукты клеток материнского организма. В некотором смысле это можно сравнить с откармливанием матки в улье группой рабочих пчел.У человека первичные половые клетки, из которых потом возникают яйцеклетки-гаметы, начинают обособляться у двухмесячного эмбриона. В возрасте 2.5 месяца они вступают в мейоз, но сразу после рождения это деление блокируется. Оно возобновляется через 14-15 лет с началом полового созревания, когда яйцеклетки раз в месяц выходят из фолликул. Но в конце второго деления мейоз снова останавливается и его блокировка снимается только при встрече со спермием. Таким образом, женский мейоз начинается в 2.5 месяца и заканчивается лишь через 20-30 и более лет, сразу после оплодотворения.
Зигота на стадии двух-восьми клеток имеет ослабленный геномный иммунитет. При изучении нестабильных инсерционных мутаций в природных популяциях дрозофилы мы обнаружили, что активация мобильного элемента, сопровождаемая мутационным переходом, происходит часто уже в первых делениях зиготы или в первых делениях первичных половых клеток. В итоге одно мутантное событие захватывает сразу клон первичных половых клеток, пул гамет становится мозаичным, и наследственные изменения в потомстве возникают пучками или кластерами, имитируя семейное наследование.
Эти эксперименты весьма важны для эпидемиологии, когда возникает вопрос о степени влияния той или иной вирусной эпидемии на генофонд потомства. Начатые еще в начале 60-х годов пионерные исследования С.М.Гершензона и Ю.Н.Александрова привели к выводу, что ДНК- и РНК-содержащие вирусы и их нуклеиновые кислоты - мощные мутагенные агенты. Попадая в клетку, они провоцируют геномный стресс, активируют систему мобильных элементов хозяина и вызывают нестабильные инсерционные мутации в группе избранных локусов, специфичных для каждого агента.
Теперь представим, что мы хотим оценить влияние на наследственную изменчивость у человека какой-либо вирусной пандемии (например, гриппа). При этом можно ожидать, что частота разного рода аномалий развития будет повышена в первом поколении у потомства, родившегося в год или спустя год после эпидемии. Оценку же частоты мутационных и вариационных изменений в половых клетках (гаметах) следует проводить во внучатом поколении.
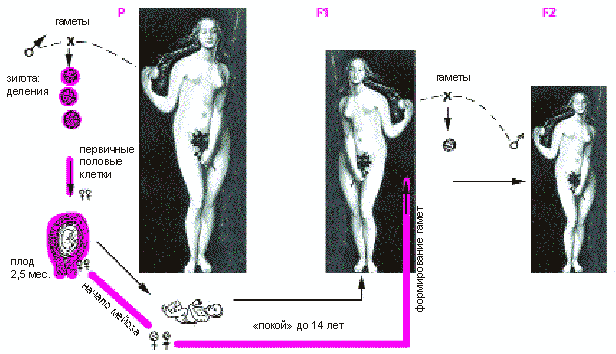
Схема оогенеза в трех последовательных женских поколениях. P - бабушка, F1 - мама, F2 - дочь.
Общий вывод состоит в том, что наследственная изменчивость у внуков может весьма зависеть от условий, в которых происходил оогенез у их бабушек! Представим женщину, которой в 2000 г. было около 25 лет, а матерью она станет в третьем тысячелетии. Оплодотворенная яйцеклетка, из которой она сама появилась на свет, начала формироваться в то время, когда ее мать была еще двухмесячным эмбрионом, т.е. где-то в середине 50-х годов ХХ в. И если в эти годы свирепствовал грипп, то его последствия должны сказаться через поколение. Для оценки последствий глобальной эпидемии на генофонд человечества надо сравнивать внучатое потомство трех групп, или когорт, - тех, у которых бабушки были беременны в год, когда разразилась эпидемия, с теми, чьи бабушки забеременели до и после пандемии (это две контрольные когорты). К сожалению, такие важные для охраны здоровья эпидемиолого-генетические сведения пока отсутствуют.
О призраках и борьбе с чудовищами
Прошло 30 лет после опытов Светлова, несложных по технике, но оригинальных по замыслу и глубоких по своим выводам. В середине 90-х годов произошел психологический перелом: резко возросло число работ в области наследственной изменчивости, в названии которых стоит слово “эпигенетический”.
Разного рода эпимутации (наследственные вариации в характере генной активности, не связанные с изменениями в тексте ДНК и носящие массовый, направленный и обратимый характер) перешли из разряда маргинальных в активно изучаемое явление. Стало очевидно, что живые системы обладают оперативной “памятью”, которая находится в непрерывном контакте со средой и использует средства природной эмбриогенетической инженерии для быстрого наследуемого перехода из одного режима функционирования в другой. Живые системы не пассивные жертвы естественного отбора, а все эволюционные формы жизни вовсе не “помарка за короткий выморочный день”, как писал Мандельштам в своем известном шедевре “Ламарк”.
Оказалось, что эпимутации сплошь и рядом можно обнаружить у обычных “классических генов”, надо только подобрать пригодную экспериментальную систему. Еще в 1906 г., за пять лет до того как Морган стал работать с дрозофилой, французский биолог-эволюционист Л.Кэно открыл у мышей менделевскую мутацию “желтое тело”. Она обладала удивительной особенностью - доминантностью по отношению к нормальной окраске (серо-коричневой) и летальностью в гомозиготе. При скрещивании гетерозиготных желтых по цвету мышей друг с другом из-за гибели гомозигот нормальные мыши появлялись в потомстве в соотношении не 3:1, а 2:1. Впоследствии оказалось, что так ведут себя многие доминантные мутации у разных организмов.
Выяснилось, что в области транскрипции одного из аллелей гена “желтое тело” внедрен мобильный элемент, напоминающий по структуре и свойствам ретровирус. В результате такой вставки ген стал подчиняться знакам пунктуации своего незваного гостя и непредсказуемо активироваться “в ненужное время и в ненужном месте”. У мутантов с инсерциями (вставками) возникают множественные дефекты (желтая окраска меха, ожирение, диабет и др.), а поведение становится нестабильным. Ненужная активность вставки в той или иной степени гасится в разных тканях за счет обратимой модификации или метилирования оснований ДНК. На уровне фенотипа проявление доминантного аллеля сильно колеблется и носит мозаичный характер. Австралийские генетики обнаружили, что у отобранных из гомогенной линии желтых самок в потомстве было больше желтых мышей, а фенотип отца -носителя мутации - не влияет на изменение окраски у потомства. Самки оказались более инерционны, и они, отобранные по фенотипу модификации ДНК, или отпечатки-импринты лучше сохранялись в оогенезе. Другие генетики нашли и чисто материнское влияние, аналогичное обнаруженному в опытах Светлова. В зависимости от диеты беременных самок выраженность мутации “желтое тело” определенным образом менялись в генотипе гетерозигот. Такое измененное состояние нестойко, но наследовалось в потомстве. Степень проявления признака коррелировала со степенью метилирования оснований ДНК во вставке.
Касаясь этих и других подобных опытов, научный обозреватель журнала “Sсience” назвал свою заметку “Был ли все же Ламарк немного прав?” Такая тактичность понятна. Во-первых, осторожность оправдана, когда приходится ревизовать то, что десятилетиями считалось прочно установленным. Во-вторых, наследование приобретенных признаков связывается не только с именем Ламарка, но и с призраком Лысенко (о последнем упоминает автор заметки). Действительно, вольно или невольно тень “мичуринской биологии” всплывает, когда обсуждается проблема наследования приобретенных признаков. И не только в России, где еще так жива память о трагедии в биологии, связанной с господством Лысенко.
Сегодня многие общепринятые положения классической генетики, которые отвергал Лысенко, стали невольно, в пику ему, считаться почти абсолютной истиной. И тем не менее, если тот или иной серьезный исследователь обнаруживал что-либо внешне созвучное взглядам Лысенко, он опасался это обнародовать, боясь остракизма со стороны научного сообщества. И даже если работа публиковалась, она сопровождалась многими оговорками и оставалась на периферии науки.
Познакомившись в 60-е годы со статьями А.А.Любищева (самого близкого друга Светлова), я пытался понять, почему он, будучи одним из самых активных самиздатных критиков лысенкоизма с 1953 по 1965 г. - его статьи и письма были собраны в книге “В защиту науки” (Л., 1990), - тем не менее не считал вопрос о наследовании приобретенных признаков окончательно решенным. Этот всеми признанный знаток эволюционной биологии указывал на незавершенность теории наследственности, на сходство наследственной и модификационной изменчивостей. Теперь мы знаем, как трудно бывает во многих случаях провести между ними границу. Любищев приводил факты массовых, быстрых и упорядоченных преобразований фенотипа в эволюции, явно необъяснимых с позиций моргановских мутаций и дарвиновского отбора. Возвысив свой голос против монополии Лысенко, Любищев выступал в защиту науки как таковой, против утвердившегося в ней аракчеевского режима. В сфере же самой науки он следовал древнему принципу: “Платон мне друг, но истина дороже”. И поэтому не скрывал своих несогласий со многими генетиками и дарвинистами. Да, Любищев оказался прав, древо жизни куда сложнее самонадеянных рассуждений.
Литература
1. Баранов В.С. и др. Геном человека и гены предрасположенности. СПб., 2000.
2. Голубовский М.Д., Чураев Р.Н. Динамическая наследственность // Природа. 1997. №4. С.16—25.
3. Чураев Р.Н. Об одной неканоническкой теории наследственности // Совр. концепции эволюц. генетики. Новосибирск, 2000. С.22—32.
4. Инге-Вечтомов С.Г. Прионы дрожжей и центральная догма молекулярной биологии // Вестн. РАН. 2000. 70(4). С.299—306.
5. Chernov Yu.O. // Mutation Res. 2001. V.488. P.39—64.
6. Хесин Р.Б. Непостоянство геномов. М., 1984.
7. Голубовский М.Д. Век генетики: эволюция идей и понятий. СПб., 2001.
8. Голубовский М.Д. // Успехи соврем. биологии. 1985. Т.100. Вып.6. С.323—339.
9. McClintock B. // Science. 1984. V.226. P.792—801.
10. Cairns J. // Nature. 1988. V.27. P.1—6.
11. Hall D. // Genetics. 1990. V.126. P.5—16
12. Shapiro J. // Science. 1995. V.268. P.373—374.
12. Бляхер Л. Я. Проблема наследования приобретенных признаков. М., 1971.
13. Landman O. // Ann. Rev.Genet. 1991. V.25. P.1-20.
14. Соколова К.Б. Развитие феногенетики в первой половине ХХ века. М., 1998.
15. Сапиенца К. // В мире науки. 1990. ?12. С.14-20.