
К 150-летию со дня рождения
Ивана Петровича Павлова
|
№ 8, 1999 г.
К 150-летию со дня рождения
|
Собаки Л.В.Крушинского
И.И. Полетаева,
доктор биологических наук
Московский государственный университет им.М.В.Ломоносова
В последние годы своей жизни Павлов пришел к выводу, что плодотворное изучение высшей нервной деятельности должно включать и анализ ее генетических основ. Это побудило его создать в Колтушах принципиально новое направление экспериментальной биологии — генетику высшей нервной деятельности. Именно тогда в первой трети ХХ в. генетика формировалась как область биологии.
В воздухе носились идеи евгеники. Неважно, что исподволь складывалось негативное официальное мнение об этой науке. Каждому здравомыслящему человеку, не только медику или биологу, было очевидно, что многие особенности организации психики человека и поведения животных имеют наследственную основу. Как подойти к экспериментальному анализу генетики поведения?
Уже существовал огромный массив двадцатилетнего опыта объективного исследования высшей нервной деятельности. По мнению Павлова, все разнообразие индивидуальных особенностей условно-рефлекторной деятельности можно было (достаточно условно) подразделить на четыре типа, соотносившиеся с классификацией Гиппократа.
Далее, было логично предположить, что принадлежность данной особи к тому или иному типу в большой степени зависит от генотипа. Для экспериментальной проверки такой гипотезы начались специальные опыты на собаках. Работа включала следующие этапы. Сначала у собак определяли тип высшей нервной деятельности (ВНД), затем формировали пары из сходных животных, а далее анализировали поведение потомков, определяя по большому ряду критериев в какой степени их условно-рефлекторная деятельность похожа или не похожа на ВНД родителей. Затем снова подбирали пары и изучали условные рефлексы у следующего поколения потомков и т.д. Такой подход позволял оценивать наследование свойств ВНД методом анализа родословных и проводить искусственный отбор по признакам поведения.
Однако так называемый “большой стандарт”, т.е. экспериментальная процедура определения типа ВНД, требовал многомесячной работы. Упрощенный вариант — “малый стандарт” — хотя и был короче, но также мало подходил для генетического анализа: он не давал возможности набрать большой экспериментальный материал. Но работа началась...
В 1936 г. Павлов скончался, а руководство Институтом экспериментальной медицины и генетическими работами в нем перешло к Л.А.Орбели. Тот в свою очередь пригласил в Колтуши в качестве консультанта по генетическим исследованиям молодого московского физиолога Л.В.Крушинского.
К этому времени выяснилось, что генетическое изучение высшей нервной деятельности собак несовместимо с длительной и очень трудоемкой процедурой определения типа ВНД по стандартам, принятым в лаборатории Павлова. Задуманные эксперименты остались незавершенными именно в связи с методическими трудностями. Как потом оказалось, перспективнее анализировать не типы ВНД, а отдельные свойства нервных процессов, например, попытаться проследить наследование сильного и слабого типов ВНД.
В 1946 г. вышла большая обзорная статья Крушинского [1]. В этой фактически первой сводке как отечественных, так и зарубежных работ по генетике поведения он привел данные по генетике “типологии” собак. В кратком виде это выглядело следующим образом: от скрещивания собак только сильного типа (шесть пометов) из полученных 20 потомков 16 оказались сильного типа, а четыре слабого; от скрещивания собак сильного типа со слабым (три помета) из полученных 12 потомков семь были сильного типа и пять слабого; от скрещивания собак слабого типа (один помет) — шесть потомков слабого типа. Полученные результаты доказывали, что различную силу типа нервной деятельности у собак можно считать наследственным свойством. Однако в этой же статье Крушинский предостерегал от более конкретных выводов о характере наследования сильного и слабого типов ВНД. Различия между животными разных типов носят количественный характер и “трудно провести демаркационную линию между ними”, что говорит о сложном наследовании такой особенности высшей нервной деятельности у собак.
Дальнейшее развитие представлений о генетических основах высшей нервной деятельности неразрывно связано в нашей стране с именем этого выдающегося ученого. Многие значительные открытия, в том числе и в физиологии, часто обусловлены, с одной стороны исторической преемственностью идей, а с другой — появлением новых “парадигм” в науке. Обе эти тенденции хорошо прослеживаются при анализе зарождения генетики поведения в СССР. Исследования Крушинского по наследованию особенностей поведения собак хотя и были логическим продолжением павловских, но принципиально отличались от того, что делали ранее. Для анализа генетико-физиологических механизмов поведения Крушинский предложил изучать наследование общей возбудимости животного.
Одно из важных достижений павловской школы объективного изучения высшей нервной деятельности — попытка количественной оценки реакции животного. Однако для такого интуитивно ясного, но вместе с тем расплывчатого понятия, как “возбудимость центральной нервной системы (ЦНС)”, не существовало сколько-нибудь общепринятых критериев и, соответственно, методов ее оценки. В него входит не только представление о некоем фоне возбудимости ЦНС, при котором протекает вся деятельность организма, в частности его поведение, но и реактивность ЦНС к внешним воздействиям. Это свойство — возбудимость животного — следовало оценивать быстро, получая надежные данные на десятках и десятках животных.
Большой опыт работы в служебном собаководстве подсказал Крушинскому, что адекватным и достаточно надежным показателем общей возбудимости ЦНС собаки может служить двигательная активность животного. Этот признак просто и быстро определяется с помощью спортивного шагомера, который крепится на теле животного. Реактивность собак на внешние воздействия Крушинский также решил измерять очень просто, характеризуя поведение собаки на появление хозяина в баллах.
В эксперимент он взял две породы собак с разным уровнем возбудимости. Гиляцких лаек, маловозбудимых собак, не обладавших пассивно-оборонительной реакцией (т.е. не обнаруживавших трусость), скрещивали со значительно более возбудимыми немецкими овчарками, у которых пассивно-оборонительная реакция также отсутствовала, но которым была свойственна злобность (т.е. активно-оборонительная реакция). Ожидать появления трусливого потомства оснований не было. Однако все вышло наоборот! У всех 25 собак, полученных от этого скрещивания (первое поколение гибридов), была резко выражена пассивно-оборонительная реакция.
Вот как описывает Крушинский поведение этих животных: “...при подходе
незнакомого лица собаки быстро убегали с поджатым хвостом и прижатыми ушами,
забивались в темный угол; зрачки у них сильно расширялись, мускулатура
тетанически напрягалась, наблюдался ясно выраженный тремор, температура
тела поднималась [2], доходя в некоторых случаях
до 40°С”.

“Выражение лица” у собак.
Такие рисунки К. Лоренц приводит как иллюстрацию
нарастания агрессивности (слева направо) и страха (сверху вниз).
И все же такой результат не очень удивил Крушинского, поскольку он, как многие собаководы, отмечал повышенную трусость гибридов немецких овчарок с волками. Волкам свойственно пассивно-оборонительное поведение, и считалось, что у гибридов с собаками оно просто в силу каких-то причин более выражено. В случае же скрещивания гиляцких лаек и немецких овчарок причиной повышенной трусливости потомства был какой-то иной фактор, поскольку родители трусливыми не были.
В то же время и для волков, и для гиляцких лаек характерна достаточно
низкая общая возбудимость, проявлявшаяся в виде невысокой двигательной
активности. Крушинский предположил, что гиляцким лайкам свойственна породная
предрасположенность к пассивно-оборонительной реакции, но она не обнаруживается
из-за низкой возбудимости. Гибриды этих собак с овчарками унаследовали
высокую возбудимость последних и пассивно-оборонительную реакцию от другого
родителя, в результате чего это свойство поведения — повышенная трусливость
— проявилось у них в полной мере. Такое объяснение подтвердилось в простом
эксперименте. Гиляцким лайкам вводили кокаин, дофаминергический агент,
который специфически активировал двигательную сферу животного. Инъекции
провоцировали у гиляцких лаек четкую пассивно-оборонительную реакцию, но
у других пород (немецкой овчарки, зырянской лайки и др.) такого эффекта
не давали. Эти данные свидетельствовали, что уровень возбудимости, тесно
коррелировавший с двигательной активностью, оказался модулятором другого
признака поведения — пассивно-оборонительной реакции.
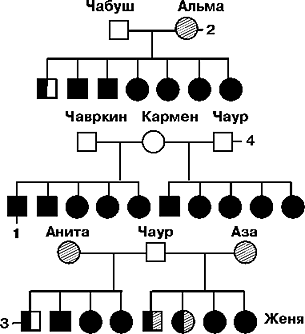
Схема скрещивания гиляцких лаек (самцы) с немецкими овчарками (самки). 1 — пассивно-оборонительная реакция (трусость), 2 — активно-оборонительная реакция (злобность), 3 — присутствие обеих реакций, 4 — отсутствие обеих реакций. (Л.В.Крушинский. 1946.)
Видно, что при скрещивании двух пород, у которых пассивно-оборонительная реакция не выражена, у потомства, как правило, проявляется трусливость.Таким образом, генетически детерминированный признак поведения может не проявляться у животных с маловозбудимым фенотипом. В то же время потомство, полученное от скрещивания таких собак с возбудимыми особями, обнаруживает подобные признаки достаточно отчетливо.
Результаты исследований, проведенных в 30—40-е годы, Крушинский
изложил в ряде оригинальных статей тех лет, а также в своей первой монографии
“Формирование поведения животных в норме и патологии”, изданной в 1962
г. Сегодня их можно найти в двухтомнике издания его избранных трудов [3].
При подготовке второго издания своей монографии “Биологические основы рассудочной деятельности” (1986), увидеть которое Крушинскому уже не довелось, он писал, что у собак уровень возбудимости ЦНС служит одним из базовых свойств, обеспечивающих выражение генетически детерминированных особенностей поведения.
Эта работа Крушинского не прошла незамеченной. В середине 40-х годов американские физиологи Дж.Скотт и Дж.Фуллер начали большой проект с подробным изучением генетики поведения собак. Их исследования также опирались на данные, полученные в собачьих питомниках, дополнялись наблюдениями за собаками, выращенными в семьях. Результаты своей многолетней работы авторы изложили в монографии, которая и по сей день продолжает оставаться одной из наиболее серьезных книг на эту тему [4].
На большом и убедительном материале Скотт и Фуллер продемонстрировали существование достаточно простой наследственной основы ряда признаков поведения. Для генетического эксперимента они выбрали две заведомо неродственные между собой породы собак — бассенджи и американские коккер-спаниели. Бассенджи (неспециализированная порода охотничьих собак ряда африканских племен) очень агрессивны и по ряду признаков напоминают диких животных. Это свойство проявляется уже у щенков бассенджи при попытках взять их в руки. Они ведут себя подобно детенышам диких животных, издают резкий визг, пытаются вырваться, укусить и т.п. Кроме того, щенки резко сопротивляются попыткам приучить их к поводку. Коккер-спаниели, напротив, — порода с низкой агрессивностью и достаточно долгой историей разведения — они легко управляемы.
Гибриды первого поколения оказались по этим признакам сходны с бассенджи. У животных второго поколения и у беккроссов (при возвратном скрещивании) картина расщепления признаков показала, что “дикость” бассенджи контролируется одним доминантным геном. Сопротивление ограничению свободы, видимо, также контролируется одним геном, но без доминирования. Возможно, что на проявление признака влияет и организм матери.
У щенков разного возраста авторы проанализировали наследование особенностей голосовых реакций в тесте на доминирование. Двух щенят в течение 10 мин заставляли бороться за обладание костью. В этой ситуации голосовая реакция щенка четко разделялась на два независимых признака. Один из них — порог провокации лая, т.е. уровень стимуляции, необходимый для проявления реакции. Он оказался высоким для бассенджи и низким — для коккер-спаниелей. Второй признак — длительность лая. Она была малой у бассенджи и большой — у коккер-спаниелей. По данным скрещиваний, низкий порог проявления лая преобладает над высоким, а частота и длительность лая наследуются по промежуточному типу. В этой работе выяснилось еще и множество других особенностей наследования признаков поведения.
Обсуждая теоретическую канву своих исследований, Скотт и Фуллер не один раз ссылаются на хорошо известную им работу Крушинского, которая помогла в осмысливании своих экспериментальных данных. Успех этой работы, как и исследования Крушинского, базировался на тщательном отборе признаков для анализа, а также на удачном подборе пород при скрещивании. Породное разведение собак хотя и не создает полной инбредности, но способствует появлению гомозиготности по многим аллелям, что и было залогом успеха работ.
Собаки — далеко не идеальный объект для изучения поведения и его нейрофизиологической, особенно генетической, основы (это было ясно еще со времен Павлова). Однако опыты с ними дали принципиально новый фактический материал, который лег в основу новых прогрессивных направлений науки. Недаром Павлов поставил в Колтушах памятник собаке, причем породистой!
И тут выстраивается далеко не случайный ряд экспериментов и экспериментаторов. Это — Павлов, который обнаружил “психическое” слюноотделение. Крушинский, эксперименты которого еще не до конца осмыслены, с наблюдениями за собаками, давшими начало учению об элементарной рассудочной деятельности. Это Скотт и Фуллер, в монографии которых уже практически современным языком изложены основные положения генетики поведения. (Генетика поведения считает себя наукой с 1960 г., т.е. с момента выхода первой обобщающей монографии.)
Данные о влиянии возбудимости на проявление и выражение генетически детерминированных признаков поведения имеют в действительности огромное общебиологическое значение. Эволюция поведения занимала многих ученых. Это очень большая тема. Здесь нам нужно упомянуть только о следующем. В соответствии с классической этологической концепцией, “единицами” поведения принято считать так называемые “фиксированные комплексы действий” (или ФКД). Набор видоспецифических ФКД образует некий поведенческий портрет вида, который, разумеется, недостаточен для полной характеристики поведения животного, но знать который необходимо иногда даже для целей таксономии.
Со времен начала этологических исследований постепенно накапливаются факты, на основании которых можно сделать некоторые умозаключения о путях формирования видоспецифических “наборов” ФКД. Вот некоторые из них.
О.Хейнрот, а затем К.Лоренц подробно изучали так называемые ритуалы ухаживания у большой группы утиных птиц. Сходство в форме таких ритуалов тем выше, чем таксономически “родственнее” друг другу сравниваемые виды. Многие виды уток скрещиваются между собой, и у гибридных особей можно наблюдать элементы ухаживания. Лоренц, например, обнаружил, что гибрид чирка и багамской шилохвости (Anas bahamensis) во время ухаживания принимает позу “голова вверх—хвост вверх”, которой нет у обоих родительских видов, но которая характерна для других видов уток. Этот феномен назвали поведенческим атавизмом.
Гибридные формы часто обладают некой комбинацией ФКД, свойственных родительским видам. Однако иногда их проявление подчиняется определенным закономерностям. Немецкий исследователь Д.Франк обратил внимание на особенность ФКД ухаживания у рыбок-меченосцев, полученных от скрещивания Xiphophorus montezumae cortezi и X.helleri. Он заметил, что гибриды ведут себя в точности как самцы X.helleri в состоянии сильного полового возбуждения и как X.montezumae при менее высоком уровне половой мотивации [5].
Немецкий физиолог А.Хаас изучал поведение шмелей. Его интересовала способность этих насекомых отыскивать гнезда (задача, традиционная для многих этологов). В эксперименте, изменяя расположение гнезда, он вызывал у шмелей аномальное возбуждение и “стресс” [6]. В таком состоянии в их поведении проявлялись новые для данного вида стереотипные элементы ФКД, которые, однако, свойственны другим видам шмелей того же рода.
Закономерности проявления некоторых видоспецифических особенностей “смертельного укуса”, характерного для всех кошачьих, исследовал известный этолог П.Лейхаузен. Он предположил, что конкретной причиной, мешающей ФКД, свойственным данному виду, проявляться в репертуаре его поведения, могут служить различия в физиологических порогах их активации. Влияние уровня возбудимости животного на проявление того или иного элемента поведения выявлялось на уровне инстинктивных реакций кошек разных видов.
Сегодня экспериментально доказано, что уровень возбудимости действительно может служить модулятором проявления элементов поведения. Этот вывод, принимаемый современными физиологами, подтверждает данные, которые более 60 лет назад получил на собаках Л.В.Крушинский. Принципиально новое объяснение, которое дал Крушинский своим результатам, сохранило свое значение и сегодня. Ведь до сих пор влияние уровня возбудимости на проявление генотипических особенностей остается малоисследованной, но по-прежнему актуальной проблемой нейрогенетики.
Современная генетика поведения — достаточно бурно развивающаяся область науки о мозге. Одно из наиболее популярных новых направлений — это анализ влияния на мозг и поведение экспериментального направленного изменения генома [7]. Такие работы ведутся преимущественно на нематодах (Caenorhab ditis clegans), дрозофиле и мышах. Изучая последствия выключения (или усиления) генов, которые экспрессируются только в мозге (нейрогенов), можно проследить закономерности его развития и работы. Уже первые итоги экспериментов на мышах показали, что эффект такого вмешательства в значительной степени определяется так называемым генетическим фоном, т.е. зависит от того, с какой линией мышей проводили опыты [8]. Иными словами, внутривидовые вариации общего строения генома могут определять конкретный результат экспрессии данного модифицированного гена.
Этот вывод еще раз возвращает нас к ранним работам по генетике поведения, о которых шла речь. Как показывает история развития идей в науке о мозге, нередко пристальное наблюдение за поведением животных в целом позволяет заметить такие детали, которые впоследствии ложатся в основу целостной интегральной концепции.
Литература
1. Крушинский Л.В. // Успехи соврем. биологии. 1946. Т.22. Вып.1. С.48—62.
2. Крушинский Л.В. // Тр. Ин-та эволюц. физиологии и патологии высш. нерв. деятельности им.И.П.Павлова. 1947. Т.1. С.39—62.
3. Крушинский Л.В. Избранные труды. М., 1991.
4. Scott P.L., Fuller J.L. Genetics and the Social Behaviour of the Dog. Chicago, 1965.
5. Frank D. The genetic basis of evolutionary changes in behaviour patterns // Ed. J.H.F.van Abeelen. The Genetics of Behaviour. Amsterdam, 1974. P.119—140.
6. Хайнд Р. Поведение животных. М., 1975.
7. Лопатина Н.Г., Пономаренко В.В. Исследование генетических основ высшей нервной деятельности // Физиология поведения. Нейробиологические закономерности. Л., 1987.
8. Craevley J.N. et al. // Psychopharmacology. 1997. V.132. P.107—124.