№ 9, 2000 г.
№ 9, 2000 г.
№ 9, 2000 г.
© М.А. ФедонкинХолодная заря животной жизни
М.А. Федонкин
Михаил Александрович Федонкин, член-корреспондент РАН,
заведующий лабораторией докембрийских организмов
Палеонтологического института РАН.Прошлое не исчезает, оно даже никогда не проходит.
У.Фолкнер
Происхождение многоклеточных животных остается одной из наиболее сложных проблем естествознания. Как и появление самой жизни, эта тайна Природы волнует поколения ученых. Такой глубокий интерес вполне объясним: многое в сверхсложном мире живого стало бы понятнее, если бы мы догадались, когда, где и почему сформировались те или иные типы физиологии, жизненных форм и экосистем.
Истоки основных биохимических механизмов функционирования живого вещества находятся в архее и раннем протерозое, отделенных от нас миллиардами лет. На протяжении большей части истории нашей планеты условия среды существенно отличались от нынешних — соответственно иными могли быть биогеохимические связи и типы физиологии. Тем не менее мы прочно связаны с далеким прошлым: многие типы биохимических реакций протекают ныне в тех же физико-химических условиях, что и в архее, с тем лишь отличием, что среды эти глубоко запрятаны в недра Земли или внутрь самих организмов — подальше от кислорода.
Высокий уровень дифференциации животного мира венда указывает на более древние корни метазоа [1]. Об этом же говорят и пока редкие находки более древних, рифейских беспозвоночных. Данные молекулярной филогенетики и модель “молекулярных часов” также свидетельствуют о довендском происхождении многоклеточных животных, хотя в зависимости от метода и класса анализируемых молекул время появления метазоа определяется довольно широко — от 1500 до 700 млн лет назад. Согласно геологической летописи, физико-химические параметры биосферы в начале и в конце этого интервала существенно различались.
“Когда” значит “где”
Вопрос о том, когда возникли первые многоклеточные, — чрезвычайно важен, поскольку он тесно связан с экологическим аспектом происхождения сложных организмов. Дело в том, что в позднем протерозое в биосфере произошли кардинальные изменения, и потому для разных моментов геологической истории нам пришлось бы строить различные сценарии происхождения животных. Таким образом, вопрос “когда?” превращается в вопрос “где и в каких условиях возникли первые животные?” Но вернемся к хронологии.
Степень дифференциации животного мира венда (от губок до членистоногих, плюс несколько групп неясного систематического положения) показывает, что корни беспозвоночных уходят в историю протерозоя. Как далеко? Находки более древних окаменелостей, которые интерпретируются как остатки беспозвоночных, пока еще очень редки, но чрезвычайно важны. В отложениях верхнего рифея России и Китая [2] обнаружены окаменелости, напоминающие современных кольчатых червей, хотя их можно сравнить и с тонкостенными хитиновыми трубками ныне живущих сцифоидных полипов. Эти прямые свидетельства животной жизни, существовавшей более 700 млн лет назад, манят палеозоологов в еще неведомые глубины прошлого Земли. Успех поиска древнейших остатков животных во многом зависит от того, насколько точно мы представляем себе условия их обитания и фоссилизации.
В попытке определить момент появления метазоа предложено несколько оригинальных подходов. Так, Дж.Валентайн полагает, что для решения этой задачи можно использовать гипотезу Дж.Боннера о том, что критерием сложности организма служит количество типов клеток в нем [3]. Действительно, современные млекопитающие имеют более 200 типов клеток, моллюски и членистоногие — более 50, плоские черви и гребневики — 20, кораллы и медузы — 7, а губки и того меньше — 5. По мнению Валентайна, наиболее ранние следы ползания, около 560 млн лет назад, оставили вендские беспозвоночные, у которых было не менее 30 типов клеток. Ретроспективная экстраполяция от современного человека (210 типов клеток) к тем относительно простым животным приводит к выводу о том, что история метазоа в докембрии очень коротка — первые многоклеточные животные появились приблизительно 680 млн лет назад.
Похожий результат получен Дж.Сепкоски, который анализировал динамику биоразнообразия в венде и кембрии [4]. Ранняя ископаемая летопись животных демонстрирует экспонентный рост разнообразия, особенно в кембрии, что также говорит о краткой истории животных. Однако “эволюционный взрыв”, который мы наблюдаем в кембрии, в огромной степени связан с появлением в животном царстве минерального скелета — шансы сохраниться в ископаемой летописи резко возросли. Многие палеонтологи полагают, что кембрийская летопись — в большей степени “взрыв изобилия окаменелостей”, чем эволюционное событие.
Резкий вызов выводам Валентайна и Сепкоски последовал со стороны молекулярной биологии. Согласно модели “молекулярных часов”, основанной на предположении о постоянной скорости изменения генома в течение длительного времени, многоклеточные животные могли возникнуть довольно рано. Так, Г.Рэй с соавторами находит весьма веские доказательства в пользу очень раннего происхождения многоклеточных животных — не позднее 1.5 млрд лет назад [5]. Сравнительный анализ семи генов, кодирующих такие белки, как глобины, показал, что эволюционные ветви беспозвоночных и хордовых разошлись около 1 млрд лет назад. Первичноротые отделились от линии, ведущей к хордовым, задолго до иглокожих — около 1200 млн лет назад. Все это говорит о продолжительной истории становления основных типов животных и противоречит гипотезе о кембрийском “эволюционном взрыве”, начавшемся 544 млн лет назад и якобы давшем начало современным типам животных. Следует заметить, что эти авторы датируют лишь время расхождения генетических линий основных типов беспозвоночных, а морфологические черты, характеризующие современных животных, например скелет или целом, могли возникнуть много позже.
О весьма длительной истории животного мира в протерозое говорят и другие специалисты, использующие метод “молекулярных часов” [6]. Явный конфликт этой гипотезы с ископаемой летописью стимулировал изучение более представительного материала: так, Ф.Аяла с коллегами, проанализировав 18 генов, кодирующих белки, заключили, что первичноротые (моллюски, кольчатые черви, членистоногие) отделились от вторичноротых (иглокожих и хордовых) около 670 млн лет назад, а хордовые и иглокожие на 70 млн лет позже. Согласно этой модели, первые многоклеточные животные появились около 700 млн лет назад, т.е. за 155 млн лет до начала кембрийского периода [7]. Единственный способ проверить надежность методов молекулярной филогенетики — вернуться к ископаемой летописи.
Слово палеонтологам
Разнообразие филогенетических деревьев, построенных зоологами и молекулярными биологами, свидетельствует об отсутствии критерия надежности реконструкций внутри каждой из упомянутых дисциплин. Вот почему взгляды неонтологов обращаются к палеонтологии, особенно к ископаемой летописи неопротерозоя (древнее 545 млн лет), где сохранились остатки наиболее древних животных. Сомнения по поводу достоверности исторических реконструкций на основе современного материала понятны: миллионы видов современных организмов составляют лишь ничтожную долю былой жизни на Земле. По оценкам палеонтологов, на протяжении более полумиллиарда лет позднейшей истории биосферы вымерло 99% видов животных. Среди них не только виды, но и целые классы и даже типы животных. Еще недавно типы животных казались “вечными таксонами”, поскольку большая часть их известна с кембрия, первого периода фанерозоя, начавшегося около 545 млн лет назад. Именно фанерозой, включающий палеозойскую, мезозойскую и кайнозойскую эры, всегда был объектом исследования классической палеонтологии.
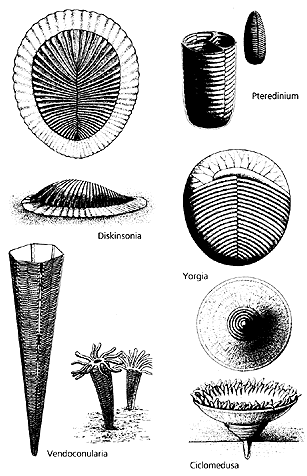
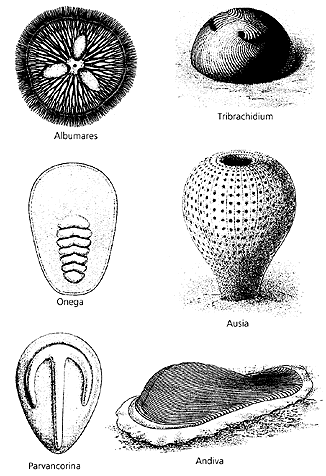
Истинным прорывом в прошлое стало открытие докембрийской фауны — крупных, подчас гигантских беспозвоночных, природа которых вызывает сейчас столько споров [8]. За последние десятилетия в составе кембрийской и более древней, вендской, фауны обнаружили животных, которых нельзя отнести ни к одному из ныне живущих типов. Несмотря на внутреннее психологическое сопротивление, первооткрывателям этих форм пришлось констатировать, что на раннем этапе животного мира существовали целые типы беспозвоночных, не нашедших продолжения. Их разнообразие могло превосходить современное, хотя видов было не так много. Как изменилась бы картина происхождения и ранней эволюции метазоа, если бы данные об эмбриональном развитии или геноме этих монстров можно было вовлечь в реконструкцию филогенеза животных?
Ряд видов вендских животных при всей их необычности представляет предковые линии кембрийских и даже ныне живущих беспозвоночных. Другие, четко обособленные группы не находят места среди современных типов. Несомненно тканевая организация, сложность строения этих организмов говорят о том, что мы имеем дело с группами весьма продвинутых, специализированных животных, которые, однако, вымерли уже в венде, т.е. более чем 545 млн лет назад. Возможно, физиологически эти беспозвоночные сформировались в условиях более древней биосферы и не смогли пережить радикальных изменений, произошедших в конце протерозоя, особенно в венде. Какие же условия царили на Земле до начала вендского периода?
Реконструкция древнейших беспозвоночных,
населявших холодноводные бассейны вендского периода
после крупнейшего Варангерского оледенения около 600 млн лет назад.Парадоксальность актуализма
или
куда девались строматолитыАктуалистические подходы к реконструкции докембрийских сред обитания зачастую приводят к неактуалистическим моделям биосферы — слишком велик интервал времени, отделяющий нас от архея и протерозоя. Примером может служить сравнение древних строматолитов и современных экосистем, контролируемых цианобактериями в себкхе или в лагунах с повышенной соленостью аридных зон. Подобные экосистемы, занимающие ныне весьма ограниченное пространство, в докембрии, напротив, были широко распространены. Этот пример ярко демонстрирует парадоксальность актуализма как метода познания геологического прошлого.
Исследователи строматолитов, тонкослоистых карбонатных структур, сформированных цианобактериальными сообществами, давно подметили резкий спад разнообразия и количества этих образований после 1 млрд лет назад. Это явление рассматривали как результат деятельности ранних животных, которые выедали бактериальные дерновинки и нарушали стабильность субстрата, необходимую для формирования строматолитов [9]. Прямые палеонтологические данные в пользу такой гипотезы отсутствуют — ни следов, ни остатков животных в строматолитовых отложениях докембрия пока не обнаружено. По чисто экологическим причинам вероятность таких находок небольшая.
Существует определенный антагонизм между строматолитами и животными: там, где много строматолитов, мало или вовсе нет животных. Даже на дне, густо заселенном животными, под островками бактериальных матов их нет из-за отсутствия кислорода. В геологической истории фанерозоя строматолиты нередко появляются в период экологических кризисов и массовых вымираний животных. Строматолиты — “знак беды”, признак дестабилизации сложных экосистем. Антагонизм устойчивых цианобактериальных сообществ и беспозвоночных заставляет нас отказаться от первой гипотезы и искать другие причины упадка строматолитов.
И.Н.Крылов, один из зачинателей палеонтологии докембрия, предположил, что роль строматолитов резко уменьшилась в результате угнетения цианобактериальных сообществ свободным кислородом, концентрация которого к концу протерозоя значительно выросла [10]. Можно сказать, что это было самоуничтожение строматолитов. Действительно, весь кислород на планете — биогенный. В архее и протерозое цианобактерии благодаря фотосинтезу произвели огромный объем кислорода. Значительная его часть ушла на окисление железа и некоторых других металлов — в результате образовались месторождения осадочных руд. После того как основная часть железа выпала в осадок, в атмосфере и гидросфере стал накапливаться свободный кислород. Таким образом, цианобактерии “отравили” атмосферу планеты продуктом жизнедеятельности и поплатились своей доминирующей ролью в мелководных морских экосистемах. По словам Крылова, это была первая глобальная экологическая катастрофа. Однако “винить” только строматолитовые бактериальные сообщества в самоуничтожении было бы неверно: в протерозое все возрастающую роль в производстве кислорода стали играть эвкариотные водоросли.
Недавно М.А.Семихатов и М.Е.Раабен опубликовали серию статей, в которых они проанализировали мировые данные по строматолитам архея и протерозоя и установили, что за период 3.8—0.545 млрд лет на фоне растущего обилия и разнообразия строматолитов было три спада [11]. Последний из них, оказавшийся фатальным, начался около 850 млн лет назад и совпал с началом длительной ледниковой эры. По мнению авторов, именно климатический фактор послужил причиной исчезновения строматолитовых сообществ.
По мере роста концентрации свободного кислорода в атмосфере протерозойской планеты уменьшалось содержание углекислого газа. В результате снижалась скорость накопления биогенных карбонатных осадков, в том числе — строматолитов. Угнетающе действовало на карбонатную биоминерализацию и длительное похолодание, возможно, связанное с уменьшением парникового эффекта атмосферы. Биосфера остывала.
Эра великих холодов
На протяжении большей части своей истории Земля не испытывала оледенений: ранняя биосфера была теплой, если не горячей. Первый ледниковый период зафиксирован геологами 2.2 млрд лет назад. За ним следовал длительный период теплой биосферы вплоть до неопротерозоя, а начиная с 750 млн лет назад в течение 200 млн лет прошла череда мощных покровных оледенений (отмечено четыре длительных ледниковых периода, но, возможно, их было больше) [12]. Масштабность этих событий потрясает. Льды достигали уровня моря даже на континентах, расположенных вблизи экватора, о чем говорят палеогеографические реконструкции, основанные на анализе остаточной намагниченности горных пород [13].
В последние годы получены дополнительные данные, показывающие поистине глобальный масштаб неопротерозойских оледенений. Изотопная летопись углерода из карбонатных отложений, накопившихся в ледниковую эру, демонстрирует негативные аномалии чудовищной амплитуды. Подобного не наблюдалось ни в течение предшествующих 1.2 млрд лет, ни в ходе всей последующей геологической истории.
Сейчас, когда гипотеза об оледеневшей Земле обрела черты научной сенсации, интенсивно обсуждается поведение биосферы. Полагают, что каждое из оледенений могло длиться от 4 млн до 30 млн лет. Тогда почти двухкилометровый слой льда покрывал континенты и мировой океан. В водах океана на миллионы лет резко сокращалась биологическая продуктивность (от фотосинтезирующего планктона и далее по пищевой цепи) — обширные льды блокировали солнечный свет. Оледенения заканчивались катастрофически быстро, когда благодаря наземному вулканизму в атмосфере повышалось содержание углекислого газа (в триста раз более его современного уровня).
Крайняя смелость этой гипотезы не означает ее несостоятельности. Она объясняет ранее непонятные явления, например такую давнюю загадку, как природа доломитов, обычно перекрывающих ледниковые отложения. Появление сразу после оледенения доломитов (CaMg(CO3)2), характерных для тепловодных карбонатных бассейнов низких широт, вызывало много споров. Полагают следующее: как только концентрация углекислого газа в атмосфере достигала критического уровня, холодная биосфера довольно быстро теплела. Океан освобождался ото льда и поглощал из атмосферы гигантские объемы углекислого газа, что вызывало осаждение доломита. В рамках этой гипотезы становится понятным и очень позднее, “возвратное” появление полосчатых железных руд, характерных для более древних эпох позднего архея и раннего протерозоя (максимум их накопления был 2.5 млрд лет назад). Покрытый льдом океан быстро терял кислород и обогащался железом за счет подводных гидротермальных источников. Освобождение океана ото льда и приток кислорода в результате возрастающей активности фотосинтезирующих организмов привели к окислению и осаждению железа.
Причины столь глубокого и длительного похолодания на планете не вполне ясны. Самая распространенная из предложенных гипотез связывает похолодание со снижением парникового эффекта атмосферы: фотосинтезирующие организмы поглотили гигантский объем углекислого газа, а захоронение биогенного углерода на обширных и стабильных континентах надолго выводило его из круговорота. Недавно появилось предположение о том, что во время позднепротерозойских оледенений наклон оси вращения Земли к плоскости ее орбиты вокруг Солнца составлял менее 36° (против нынешних 66.5°) и потому самыми холодными оказались экваториальные районы, а более теплыми — полярные области. Такая гипотеза требует объяснения механизма столь быстрых и периодических изменений наклона оси [14]. Как видим, споры идут лишь о масштабности оледенений, а в том, что в интервале 750—545 млн лет назад было несколько ледниковых периодов, сомнений нет.
Массовому расселению вендской фауны непосредственно предшествовало самое обширное Варангерское оледенение. Какую роль сыграли оледенения в эволюции многоклеточных организмов? Как они смогли пережить эру великих холодов? Прежде чем рассматривать этот вопрос, обратимся к наиболее древней части ископаемой летописи животных — вендской фауне.
Признаки холодноводности вендской фауны
Подавляющее большинство местонахождений вендских животных обнаружено в обломочных породах (песчанике, алевролите, глине), которые накапливались в условиях умеренного или даже холодного климата. Известны находки, хотя и редких остатков метазоа, даже между слоями тиллитов — ледниковых отложений. Вендские морские бассейны, населенные животными, находились вне карбонатного пояса планеты. Сейчас этот пояс располагается в низких широтах с теплым климатом и отличается высоким биоразнообразием. А в венде, как и на протяжении почти всей предшествующей истории, мелководные карбонатные бассейны контролировались цианобактериями. Об относительной холодноводности вендских метазоа говорит и массовая сохранность их остатков — низкие температуры сдерживают распад органического вещества. Экспериментально доказано, что температурный оптимум многих бактериальных ферментов, ответственных за разложение сложных органических соединений, намного выше температуры морских вод арктических бассейнов.
Есть и другие признаки, по которым вендская фауна схожа с той, что населяет современные холодные воды. Прежде всего, это преобладание мягкотелых форм и отсутствие минерального (карбонатного) скелета. Растворимость таких минералов, как арагонит и кальцит, формирующих раковины моллюсков, скелет кораллов, значительно выше в холодных водах. Соответственно энергетические затраты на постройку карбонатного скелета в холодных водах выше, чем в теплых. Вот почему животные тропических бассейнов отличаются более массивным скелетом от родственных форм холодноводных обитаний.
Замечена некоторая корреляция между присутствием эндосимбиотических водорослей зооксантелл и способностью строить карбонатный скелет у ныне живущих кораллов. В умеренных и высоких широтах эндосимбиоз одноклеточных водорослей с животными не столь распространен, как в тропиках, — сезонность, а тем более полярная ночь делают их присутствие малоэффективным.
Изучение биотических и экологических характеристик современных холодных вод и вендских палеобассейнов показывает ряд общих черт. Среди них: обильная жизнь в море по сравнению с сушей; высокая биомасса бентоса; преобладание мягкотелых в бентосных сообществах; множество прикрепленных форм среди бентосных беспозвоночных; крупные размеры тела; сравнительно низкое биотическое разнообразие; уникально короткие трофические цепи; большое количество аберрантных форм; большая доля кишечнополостных среди хищников; высокая плотность популяций только некоторых видов; низкая физическая активность роющих форм; незначительная роль бактерий; низкая скорость биологической деструкции и другие особенности. Неравномерное распределение остатков фауны (от весьма обильных до редких) в однотипных отложениях, возможно, отражает сезонность размножения и расселения вендской фауны.
Плюсы и минусы холодных вод
Современные холодные бассейны характеризуются силикокластической седиментацией и отсутствием карбонатных осадков. Активная вертикальная циркуляция вод, высокое содержание фосфатов, нитратов, растворенного органического вещества создают условия для очень высокой продуктивности первичных продуцентов — фитопланктона, начального звена большинства пищевых цепей. Это компенсирует недостаток света из-за сезонной изменчивости светового режима, обширной облачности и низкой прозрачности воды. Высокая растворимость кислорода служит важнейшим фактором для всех эвкариот, особенно для животных (морская вода при 0°С поглощает в 1.6 раза больше кислорода, чем при 20°С). Большинство биологических процессов (питание, рост, размножение, расселение и смертность) определяются сезонными флуктуациями светового режима через фотосинтезирующий фитопланктон. В холодных регионах мы видим резкий контраст между богатой морской экосистемой и скудной жизнью на суше. Не так ли было и в протерозое до того, как сушу заселили высшие растения? Холодноводные бассейны отличаются высокой биомассой и продуктивностью биоты, очень короткими трофическими цепями (фитопланктон-криль-кит или фитопланктон-донные детритофаги), низким биоразнообразием, малой стабильностью биоценозов, преобладанием травоядных планктотрофов среди планктонных беспозвоночных и форм с прямым развитием (без стадии пелагической личинки), небольшой долей хищников в планктоне (среди них много медуз и гребневиков). Для многих видов характерны высокая продолжительность жизни индивида и неограниченный, хотя и медленный, рост, низкие скорость смены поколений и репродуктивный потенциал (яйца относительно крупные, богатые желтком, но немногочисленные), медленное распространение. Небольшие скорости метаболизма, измеренные по поглощению кислорода, и невысокая физическая активность связаны с низкими температурами. Стабильность донных биоценозов относительно мала, при том что некоторые виды образуют весьма плотные поселения с высокой биомассой — результат длительного роста, позднего созревания и слабого давления хищников. Повышенная вязкость холодной воды влияет на ее структуру, подвижность флюидов в теле животного, эффективность фильтрующих систем. Холодная вода дольше держит взвесь, в том числе фитопланктон — отсюда ее большая мутность.
Сходство современных холодноводных экосистем с вендскими сообществами направляет поиск еще более ранних животных не в теплые тропические воды древних морей, а в прохладные воды более высоких широт. Есть и другой резон искать в протерозое первичные биотопы древнейших животных вне карбонатного пояса Земли. Дело в том, что теплые мелководные бассейны, в частности обширные моря рифея, контролировались прокариотами вплоть до конца венда. А.В.Вологдин, один из пионеров палеонтологии докембрия, полагал, что древние цианобактерии, как и современные, умели защищать себя ядами, угнетающими рост и размножение эвкариот, а в ряде случаев приводящими к их гибели. Так что заселить тепловодные бассейны высшим организмам было непросто.
Из холода в тепло — завоевание тропиков
Сезонность питания, характерную для высоких широт, можно рассматривать как фактор отбора в пользу форм с большей массой. Так называемая резервная биомасса нужна, чтобы переживать неблагоприятные времена. Однако рост и размеры тела ограничены обменными процессами — прежде всего дыханием. Развитие гетеротрофии и эффективных способов сбора пищи могли способствовать созданию резервной биомассы и больших размеров тела только при достаточной концентрации кислорода в воде. Такое преимущество давали холодноводные бассейны.
Путь из холодных, богатых кислородом, вод в теплые стал возможным, когда в атмосфере резко выросло содержание свободного кислорода. По данным изотопного анализа углерода из позднего докембрия, это событие произошло в самом конце протерозоя [15].
Первую попытку животных колонизовать тепловодный карбонатный бассейн мы наблюдаем на примере отложений Оленекского поднятия (север Якутии). Когда по окончании Варангерского оледенения морские воды начали затапливать континент, животные быстро заняли теплые мелководья. Вендские беспозвоночные довольно долго “удерживали свои позиции” — остатки мягкотелых, преимущественно кишечнополостных, в изобилии встречаются в битуминозных тонкослоистых известняках хатыспытской свиты мощностью почти 100 м. Трудно сказать точно, сколько длился этот эпизод, но цианобактериальные сообщества надолго “взяли реванш” — мощность строматолитовых доломитов залегающей выше туркутской свиты более 200 м. Судя по современным аналогам, строматолиты растут крайне медленно. Лишь в самом конце венда и особенно в начале кембрия сообщества животных смогли вернуться в свободные от строматолитов обитания.
Причины массовой биоминерализации
В свете описанного сценария становится понятной загадочная массовая биоминерализация в разных группах животного царства в начале кембрия. Животные появились в относительно холодных водах вне карбонатного пояса планеты, который контролировали прокариоты [16]. Эра великих оледенений (криоген) давала большее преимущество именно эвкариотам, в том числе животным, хотя это было время их трудной эволюции. В ту холодную пору площади карбонатных бассейнов и ареалы прокариотных сообществ резко сократились. По окончании криогена животные смогли заселить тепловодные бассейны карбонатного пояса, а карбонатные постройки цианобактерий постепенно замещались рифами. Это обстоятельство резко ускорило эволюционные процессы, в частности — на основе сформированного минерального скелета.
Рост разнообразия эвкариот в целом способствовал удлинению пищевых цепей. Однако в тканях животных, находящихся на вершине трофической пирамиды, могли накапливаться высокие концентрации ряда элементов, в частности Ca, P, Si. Возникла необходимость их выведения или детоксикации. У части беспозвоночных появилась возможность строить минеральный скелет вследствие детоксикации в условиях тепловодных местообитаний, где растворимость ряда биоминералов ниже, а эффективность ответственных за минерализацию ферментов выше, чем в холодных водах. В становлении биоминерализации животных не исключена и некоторая роль симбиотических микроорганизмов. Специалистам по кораллам известна замечательная закономерность: виды, имеющие симбиотические водоросли (зооксантеллы), формируют прочный массивный скелет, и наоборот, — у видов без симбиотических водорослей скелет минерализован слабо или минерализация вовсе отсутствует. Холодноводная фауна протерозоя, обитавшая в высоких широтах, могла не иметь устойчивого симбиоза с одноклеточными водорослями по причине краткого светового дня зимой. Колонизация тропиков и гарантированный световой день сделали этот симбиоз более эффективным в двух аспектах — снабжении кислородом хозяина и расширении возможностей биоминерализации.
Заключение и прогноз
Нам точно не известно, когда возникли первые животные. Разные подходы определенно указывают, что это событие произошло до начала венда, т.е. не позднее 700 млн лет назад. Ранняя эволюция животных протекала в условиях холодной биосферы. Имея в виду, что температурный оптимум эвкариот в среднем ниже, чем для прокариот, можно предположить, что холодноводные условия в большей степени угнетали бактериальные сообщества, чем высших организмов. С тем же знаком было влияние растущей концентрации кислорода в атмосфере. Фотосинтезирующие эвкариотные водоросли все прочнее занимали место цианобактерий. На протяжении позднего протерозоя области доминирования микробной биоты неуклонно сокращались.
Череда ледниковых периодов неопротерозоя дала шанс для длительной, хотя и драматичной эволюции ранних животных. До конца протерозоя многие группы животных все еще обитали в прохладных бассейнах. Высшие организмы, пережившие 200 млн лет преимущественно холодной биосферы, смогли бросить вызов архаичной бактериальной биоте и с начала кембрия заселили теплые бассейны карбонатного пояса планеты. Это событие имело колоссальное значение для всей последующей эволюции животной жизни. Просматривая этот сценарий в надежде найти истоки высокоорганизованной жизни, мы неизбежно придем к холодным бассейнам, где бактерии не могли играть определяющей роли. Именно туда направляет поиск остатков более древних, довендских животных гипотеза “холодной колыбели”. На ранней Земле такие области могли быть относительно небольшими, но с наступлением ледниковой эры неопротерозоя ареалы эвкариот существенно увеличились. Как ни парадоксально это звучит, но “что плохо для жизни, то хорошо для эволюции”. Спасибо льдам протерозоя!
Исследование выполнено при поддержке Российского фонда фундаментальных исследований.
Гранты 99-05-64547 и 99-05-65004.Литература
1 Соколов Б.С. Вендский период в истории Земли // Природа. 1984. №12. С.3—18; Он же. Очерки становления венда. М., 1998.
2 Гниловская М.Б. // ДАН. 1998. Т.359. №3. С.369—372; Sun Weiguo, Wang Guixiang and Zhou Benhe // Precambrian Research. 1986. V.31. P.377—403.
3 Valentine J.W. The Cambrian explosion / Bengtson S. Early Life on Earth. Nobel Symposium №84. N.-Y., 1994. P.401—411.
4 Sepkoski J.J.Jr. Proterozoic-Early Cambrian diversification of metazoans and metaphytes // The Proterozoic Biosphere. A Multidisciplinary Approach / Eds J.W.Schopf, C.Klein. Cambridge, 1992. P.553—561.
5 Wray G. A., Levinton J.S., Shapiro L.H. // Science. 1996. V.274. P.568—573.
6 Bromham L., Rambaut A., Fortey R., Cooper A., Penny D. // Proc. Natl. Acad. Sci. U.S.A. 1998. V.95. №21. P.12386—12389.
7 Ayala F. J., Rzhetsky A., Ayala F.J. // Ibid. 1998. V.95. №2. P.606—611.
8 Федонкин М.А. Загадки вендской фауны // Природа. 1989. №8. С.59—72.
9 Awramik S.M. // Science. 1971. V.174. P.825—827.
10 Крылов И.Н. Древнейшие следы жизни // Природа. 1985. №9. С.68—76.
11 Семихатов М. А., Раабен М. Е. // Стратиграфия. Геол. корреляция. 1994. Т.2. №6. С.10—32; Они же. // Там же. 1996. Т.4. №1. С.26—54.
12 Kaufman A.J., Knoll A.H., Narbonne G.M. // Proc. Natl. Acad. Sci. U.S.A. 1997. V.94. №13. P.6600—6605; Kennedy M.J., Runnegar B., Prave A.R., Hoffmann K.-H., Arthur M.A. // Geology. 1998. V.26. №12. P.1057—1192.
13 Чумаков Н.М. Климатический парадокс позднего докембрия // Природа. 1992. №4. С.34—41.
14 Williams D.M. et al. // Nature. 1998. V.396. P.453.
15 Knoll A.H., Carrol S.B. // Science. 1999. V.284. №5423. P.2045—2220.
16 Fedonkin M.A. // Paleontological Journ. 1996. V.30. №6. P.669—673.
 Август 2000 |