№ 9, 2000 г.
№ 9, 2000 г.
№ 9, 2000 г.
© И.С. Даревский, В.В. Гречко, Л.A. Куприянова
Ящерицы, размножающиеся без самцов
И.С. Даревский, В.В. Гречко, Л.A. Куприянова
Илья Сергеевич Даревский, член-корреспондент РАН, доктор биологических наук,
гл. научн. cотр. лаборатории орнитологии и герпетологии Зоологического института РАН.Верната Викторовна Гречко, кандидат биологических наук,
ст. научн. сотр. лаборатории эволюции эукариотических геномов Института молекулярной биологии РАН.Лариса Андреевна Куприянова, кандидат биологических наук,
ст. научн. сотр. лаборатории орнитологии и герпетологии Зоологического института РАН.
С детства мы умеем различать кота и кошку, курицу и петуха, т.е. знаем о существовании мужского и женского полов, причем не только в царстве животных, но и растений. Тем не менее укоренившееся в сознании людей представление о неизбежной двуполости всех живых существ не соответствует действительности. В настоящее время известно большое число видов растений и животных, у которых мужских особей нет вовсе, а если они периодически и появляются, то лишь на короткое время. Отсутствие мужского пола не сказывается, однако, на судьбе однополых видов — самки регулярно размножаются без участия самцов и, следовательно, без оплодотворения.
Долгое время считалось, что к однополому размножению (партеногенезу) среди животных способны лишь некоторые беспозвоночные — различные черви, моллюски и насекомые. Поэтому настоящей сенсацией стало сделанное нами в конце пятидесятых годов открытие однополого размножения у таких высокоорганизованных позвоночных, как пресмыкающиеся, а именно — у некоторых видов кавказских скальных ящериц, относящихся к роду Lacerta [1]. Эти ящерицы принадлежат к числу самых обычных и широко распространенных позвоночных горного Кавказа. На всем протяжении от Предкавказья и Дагестана до юга Армении и Азербайджана, от Черноморского побережья до ледников Главного Кавказского хребта едва ли отыщется ущелье или обнажение скал, где не встречались бы эти небольшие юркие пресмыкающиеся.
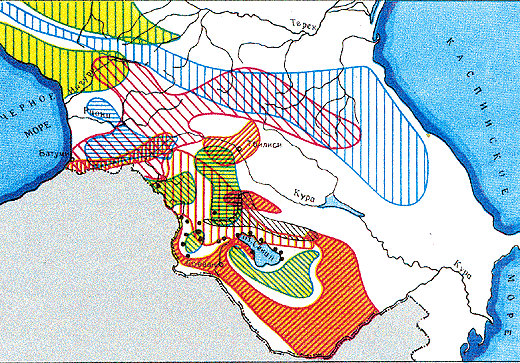
Современные ареалы скальных ящериц в пределах Кавказа.
Видны более или менее обширные зоны совмещения ареалов различных видов.Справа - маркировка ареалов сверху вниз:
Двуполые виды собственно скальная ящерица
кавказская ящерица
с подвидами
грузинская ящерица
с подвидами
аджарская ящерица
краснобрюхая ящерица
ящерица Валентина
азербайджанская ящерица с подвидами
куринская ящерицаПартеногенетические виды армянская ящерица
ящерица Даля
ящерица Ростомбекова
белобрюхая ящерицаХарактерная особенность скальных ящериц — ярко выраженная полиморфность: одни виды окрашены одноцветно и скромно и характеризуются большим постоянством морфологических признаков, другие отличаются разнообразием и яркостью расцветки, третьи же сочетают признаки, свойственные одновременно нескольким разным видам. В пределах своего обширного ареала, охватывающего, помимо Кавказа, горный Крым, Малую Азию и северную часть Ирана, они образуют значительное число видов, подвидов и популяций, длительное время объединявшихся под сборным названием Lacerta saxicola, которое дал им известный русский зоолог Э.А. Эверсманн (1794—1860).
Разобраться во всем этом многообразии форм, отыскать надежные и стойкие критерии их разграничения — вот задача, которая с давних пор привлекала к себе внимание отечественных и зарубежных систематиков. Однако несмотря на многочисленные исследования, сделать это оказалось совсем не просто. Очень скоро выяснилось, что дальнейшее накопление и без того обширного коллекционного материала расширяет спектр полиморфизма и не только не способствует решению задачи, но, напротив, все более ее усложняет. Трудности особенно возросли, когда выяснилось, что различные виды скальных ящериц легко друг с другом скрещиваются, образуя морфологически трудно различимые гибриды. Стало ясно, что выход из создавшегося положения заключается не в поиске более тонких диагностических признаков, а в признании того, что полиморфизм скальных ящериц — нормальная форма их существования. В связи с этим весь обширный накопившийся материал необходимо было пересмотреть с принципиально иных теоретических и методологических позиций. Потребовалась обработка не только мертвого коллекционного материала, но и изучение живых ящериц в их естественной обстановке, ибо лишь таким путем могут быть выявлены действительные отношения, существующие между отдельными популяциями вида в природе.
В результате многолетних исследований нам удалось выделить и описать внутри группы L.saxicola еще как минимум 18 самостоятельных видовых форм на основании анализа их морфологических признаков и зоогеографии [2]. Именно в процессе этих полевых и лабораторных исследований было обнаружено, что семь видов скальных ящериц практически лишены самцов и размножаются без их участия посредством партеногенеза. Встреченное поначалу с недоверием, наше открытие впоследствии было поддержано отечественными и зарубежными зоологами. По понятной причине изучение именно таких однополых ящериц легло в основу наших комплексных исследований, поддержанных грантами РФФИ в 1993 и 1996 гг.
В первую очередь были отобраны четыре широко распространенных двуполых вида — ящерица Валентина (L.valentini), азербайджанская (L.raddei), куринская (L.portschinskii) и аджарская (L.mixta), а также четыре партеногенетических — ящерицы Даля (L.dahli), Ростомбекова (L.rostombekovi), армянская (L.armeniaca) и белобрюхая (L.unisexualis), встречающиеся, в основном, в Армении и прилежащих районах Грузии и Азербайджана. Партеногенетические виды морфологически оказались очень сходны со своими двуполыми родственниками, и естественно возник вопрос: каким образом они образовались? Возможно, партеногенетические виды среди скальных ящериц появились в результате межвидовой гибридизации. Это предположение возникло, когда выяснилось, что они обладают некоторыми промежуточными признаками окраски и чешуйчатого покрова, и выглядело достаточно убедительно, поскольку ранее, в опытах на шелковичных червях, уже была известна потенциальная склонность гибридов к однополому размножению [3].
По техническим причинам рисунок будет вставлен позже.
Схема гибридогенного видообразования
четырех партеногенетических (2) видов скальных ящериц
в результате скрещивания в различных комбинациях
четырех родительских (1) двуполых форм.Для проверки своей гипотезы мы использовали некоторые современные методы молекулярной систематики, в частности аллозимный анализ, основанный на сравнении электрофоретической подвижности видоспецифических белков восьми перечисленных двуполых и партеногенетических видов [4]. Анализ по нескольким информативным локусам выявил в электрофореграммах всех четырех партеногенетических видов активные аллели двух разных бисексуальных, предположительно родительских форм, что свидетельствует об их происхождении в результате межвидовой гибридизации. Примечательно, что спаривание в разных комбинациях одних и тех же родительских видов привело к формированию различных партеногенетических форм. Однако это требовало дальнейшего уточнения в связи с тем, что в пределах Кавказа обитают еще несколько двуполых видов — собственно скальная ящерица (L.saxicola), кавказская (L.сaucasica), дагестанская (L.daghestanica), грузинская (L.rudis), краснобрюхая (L.parvula) и некоторые другие ящерицы. Если судить по морфологическим признакам, то они также могли претендовать на участие в гибридогенном образовании четырех однополых форм, несмотря на то, что в настоящее время географически они значительно удалены друг от друга.
Для того чтобы разобраться в этом, мы использовали некоторые другие современные методы молекулярной систематики, в частности анализ ядерной и митохондриальной ДНК. Особенно важную роль сыграл предложенный нами метод рестриктазного картирования нуклеотидных повторов геномной ДНК, получивший название “таксонопринта”. Дело в том, что размер, структура последовательностей и число повторов обладают видо- и родоспецифическими свойствами, которые могут быть использованы как молекулярные признаки этих таксонов. Эта характеристика повторов ДНК одинакова у каждого индивида популяции (это было показано на обширном материале, полученном не только на ящерицах, но и на других группах животных — насекомых, рыбах, млекопитающих) и представляет собой его как бы “паспорт”. Более того, близкие популяции одного вида не различаются по этой характеристике и потому могут быть объединены в один вид, независимо от степени характеризующего их полиморфизма. Разные же виды обладают специфическими картинами повторов, которые, однако, сохраняют некоторые общие признаки, что позволяет заключить их в рамки одного рода. У метода таксонопринта немало и других достоинств, например с его помощью удалось выявить родительские виды для некоторых партеногенетических. Выяснилось, что из большого числа популяций широко распространенного в Закавказье двуполого вида азербайджанской ящерицы только одна из них может претендовать на роль родительской по отношению к партеногенетической ящерице Ростомбекова [5].
Не менее интересные данные были получены с помощью метода полимеразной цепной реакции с использованием единичного случайного праймера (RAPD), который широко используется в систематических исследованиях для характеристики внутри- и межпопуляционного полиморфизма, а также для выявления различий между популяциями различных видов. Оказалось, что отдельные партеногенетические таксоны содержат в своем геноме локусы ДНК двух бисексуальных видов (т.е. по своему происхождению эти таксоны гибридны) и, более того, не имеют своих собственных видоспецифичных локусов, которые были бы характерны только для них. Все особи партеногенетической популяции обладают совершенно одинаковыми картинами RAPD-маркеров. Это свидетельствует как о чрезвычайной устойчивости генома партеногенетических видов, так и об их относительно недавнем с эволюционной точки зрения происхождении [6].
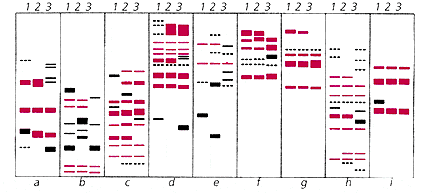
Сравнение полиморфизма RAPD с использованием различных праймеров (a—i)
у гибридогенных партеногенетических видов скальных ящериц L.rostombekovi (1), L.dahli (3)
и одного из родительских, по отношению к ним, двуполых видов L.portschinskii (2).
Локусы ДНК, заимствованные обоими однополыми видами
от их двуполой “отцовской” формы, выделены цветом.Оба метода однозначно указывают на гибридное происхождение четырех партеногенетических форм скальных ящериц. Аллозимный анализ партеногенетических ящериц выявил в той или иной мере свойственную им генетическую неоднородность, которая проявляется в так называемой поликлонии. Например, у однополой ящерицы Даля обнаружено не менее пяти клонов, у армянской и белобрюхой — по три клона, тогда как ящерица Ростомбекова (в пределах изученной части ее ареала) — моноклональна [7]. Это может свидетельствовать о происхождении таких видов в результате не одного, а нескольких актов межвидовой гибридизации, видимо, имевших место в разное время и в различных частях их ареалов.
Цитогенетический анализ двуполых и партеногенетических видов скальных ящериц поначалу нас разочаровал, так как по кариотипу все они оказались очень сходны и имеют стандартный диплоидный набор гомологичных акроцентрических хромосом (n = 38). Ситуация явно неудачная, так как у некоторых других гибридогенных партеногенетических видов ящериц хромосомные наборы гетероморфны, т.е. состоят из различных по морфологии хромосом, заимствованных от кариотипически различающихся родительских пар. Например, если один из родительских видов характеризуется метацентрическими хромосомами (с перетяжкой в средней части), а другой — субметацентрическими (с перетяжкой, сдвинутой к одному из концов), то диплоидный кариотип порожденного от их гибридизации вида включает гаплоидные наборы метацентрических и субметацентрических хромосом, полученных от каждого из родителей. Именно так обстоит дело у многих американских однополых ящериц из рода Cnemidophorus. У “наших” же партеногенетических видов скальных ящериц хромосомные наборы (за одним исключением, на котором мы остановимся позже) — гомологичны.
И все же нам удалось обнаружить некоторые хромосомные маркеры и подтвердить гибридную природу партеногенетических видов Lacerta. Выяснилось, например, что хромосомы некоторых из них характеризуются разным распределением специфически окрашенных гетерохромативных районов. Так, у одного из родительских видов в хромосоме “А” первой пары такой окрашенный район располагается на верхнем ее конце, тогда как у второго из родителей — лежит ближе к центру или отсутствует вовсе. Соответственно у партеногенетического вида диплоидный набор имеет по одной разноокрашенной хромосоме, заимствованной от обоих родителей, что указывает на имевшую место межвидовую гибридизацию. Другой стойкий маркерный признак кариотипов скальных ящериц проявляется в виде присутствия мелкой (редуцированной) половой хромосомы (типа W), которую все партеногенетические виды унаследовали от разных материнских видов, тогда как вторая половая хромосома (Z) досталась им от отцов. Другими словами, у скальных ящериц носитель гетероморфных половых хромосом — не мужской пол (по типу XY), а женский (по типу WZ), а по типу ZZ у них гомогаметны самцы [8]. Мы уже говорили, что скальные ящерицы легко вступают в естественную межвидовую гибридизацию, причем по понятной причине для нас особенно были интересны гибриды между партеногенетическими самками, спаривающимися с самцами обитающих совместно бисексуальных форм.
Говоря о гибридизации, напомним, хотя бы в общих чертах, о роли оплодотворения в процессе размножения у животных. Не вникая в цитогенетические подробности, можно сказать, что биологический смысл этого процесса — происходящее при оплодотворении слияние мужской половой клетки (сперматозоида) с женской половой клеткой (яйцеклеткой). От всех остальных (соматических) клеток организма обе эти половые клетки отличаются прежде всего редуцированным (уменьшенным вдвое) числом хромосом. Так, если в соматических клетках скальных ящериц имеется, как мы помним, по 38 хромосом (2n = 38), то в каждой из половых клеток их 19 (n = 19). Нормальное удвоенное число хромосом называют при этом диплоидным, а уменьшенное вдвое — гаплоидным. При слиянии мужской и женской половых клеток (гамет) число хромосом в оплодотворенном яйце (зиготе) увеличивается до диплоидного, и развивающийся из него зародыш, как и оба родителя, вновь имеют 2n = 38. Это довольно сложный процесс, но в данном случае достаточно знать, что оплодотворенное яйцо неизменно заимствует равное число хромосом от отца и матери. Однако так происходит лишь при нормальном оплодотворении с участием обоих полов.
У партеногенетических же видов слияние мужской и женской половых клеток не происходит, тем не менее развивающийся из яйца зародыш также обладает удвоенным числом хромосом. Это, противоречивое на первый взгляд, обстоятельство объясняется тем, что при однополом размножении недостающий гаплоидный набор образуется не за счет половинного числа хромосом, привносимых сперматозоидом, а восстанавливается в результате сложных цитогенетических преобразований, протекающих внутри самой яйцеклетки. В зависимости от типа партеногенеза характер таких преобразований бывает различен, но в результате всегда восстанавливается удвоенный набор хромосом, с той лишь существенной разницей, что теперь он имеет уже не смешанное материнское и отцовское происхождение, а обе составляющие его половины — производные одной и той же женской половой клетки. Понятно, что образующийся при этом зародыш лишен отцовской генетической информации, со всеми вытекающими из этого последствиями, сказывающимися, в частности, на морфологических признаках развивающегося партеногенетического потомства.
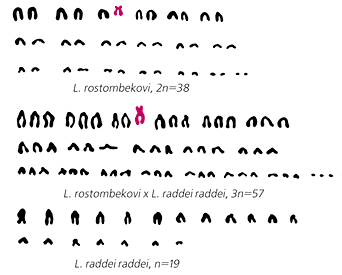
Диплоидный набор хромосом партеногенетической самки L.rostombekovi,
гаплоидный набор самца двуполого вида L.raddei
и триплоидный набор стерильного гибрида между ними.
Имеющиеся в наборах материнской формы и гибрида
непарные субметацентрические хромосомы отмечены цветом.Представим себе теперь, что же происходит при спаривании партеногенетической самки с самцом одного из встречающихся совместно двуполых видов. В этом случае к партеногенетически удвоенному чисто женскому хромосомному набору вместе со сперматозоидом добавится третий мужской гаплоидный набор, в результате чего зигота и развивающийся из нее зародыш станут триплоидными — 3n = 57. Такие аллотриплоидные (происходящие от спаривания разных видов) гибриды действительно встречаются в природе и составляют иногда до 10—12% всех особей в смешанной популяции. Как и следовало ожидать, они обладают признаками отцовского и материнского видов, причем материнские признаки, в соответствии с двойной дозой женского генома, у них преобладают над отцовскими. Добавим, что у триплоидных гибридов резко выражен так называемый гетерозис, проявляющийся, в частности, в заметном возрастании размеров тела и головы. Не удивительно поэтому, что нахождение в природе столь необычных ящериц приводило в отчаянье систематиков, которые, не ведая об истинной их природе, безуспешно пытались привязать их то к одному, то к другому из уже известных видов или, более того, выделять в качестве самостоятельных видовых форм. Естественно, триплоидный кариотип гибридов содержит два хромосомных набора, полученных от самки и один — от спарившегося с ней самца. Это особенно ярко демонстрируется в случае, когда матерью становится ящерица Ростомбекова, специфический кариотип которой содержит одну субметацентрическую хромосому, присутствующую также в хромосомном наборе гибрида.
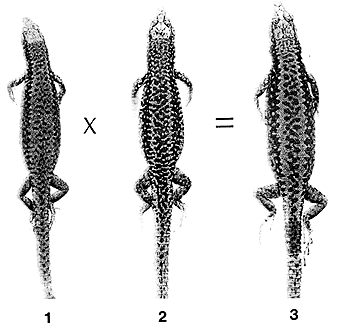
Партеногенетическая самка L.armeniaca (1),
самец двуполого вида L.valentini (2)
и образующийся в результате их скрещивания стерильный триплоидный гибрид (3).
Обращают на себя внимание более крупные, чем у родителей, размеры тела последнего.Среди триплоидных гибридов встречаются как самки, так и мужские особи, причем первые из них лишены яичников и полностью стерильны, а плодовитость вторых если и проявляется (в их семенниках обнаружены жизнеспособные сперматозоиды), то значительно понижена. Триплоидные особи, как и все нечетно-полиплоидные организмы, в случае когда они плодовиты, к бисексуальному размножению не способны и могут производить себе подобных лишь посредством партеногенеза. Большая часть партеновидов Cnemidophorus имеет именно такую природу.
Среди различных групп ящериц в настоящее время известны многие плодовитые триплоидные партеногенетические виды, в том числе и описанные нами некоторые вьетнамские гекконы и агамы. Что же касается скальных ящериц, то их триплоидные самки, как уже говорилось, представляют собой эволюционный тупик, тогда как у самцов есть некоторая эволюционная перспектива, связанная с теоретической возможностью продуктивного спаривания с самками как двуполых, так и партеногенетических видов. Пока это лишь плод наших размышлений и требует специального изучения, чем мы и планируем заняться в дальнейшем.
Очевидное преимущество однополого размножения у скальных ящериц заключается в том, что благодаря ему происходит удвоение темпов размножения вида, поскольку все особи в популяции представлены только самками. Действительно, если иметь в виду, что у бисексуальных видов по крайней мере половина особей — не оставляющие потомства самцы, то преимущества, связанные с заменой их самками, становятся очевидны. Таким образом, партеногенетическое размножение постоянно способствует поддержанию высокой численности вида, что особенно важно, если в силу тех или иных причин, как это наблюдается иногда в действительности, она может подвергаться значительным колебаниям. Интересно, что если в популяциях двуполых видов, особенно в период размножения, наблюдаются необычайная суета и оживление, вызванные дерущимися друг с другом и преследующими самок самцами, то в плотных поселениях партеногенетических самок обычно царят “мир и благоволение” и их можно наблюдать большими группами, бок о бок мирно обогревающимися под солнцем.

Группа самок партеногенетического вида L.unisexualis,
обогревающихся под солнцем в естественных условиях.Принято считать, что подавляющее большинство всех видов современных животных исходно образовалось в процессе медленной дивергентной эволюции, в соответствии с классической теорией Дарвина. Что же касается партеногенетических видов скальных ящериц, то, как нам удалось показать это с помощью различных методов, они образовались принципиально иным путем: в результате не разделения, а напротив — схождения филетических линий, т.е. благодаря естественной межвидовой гибридизации. Схематически этот процесс можно представить в виде характерной сети из сходящихся и перекрещивающихся филумов, откуда происходит и название самого термина “сетчатая”, или “ретикулярная” (у англоязычных авторов), эволюция.
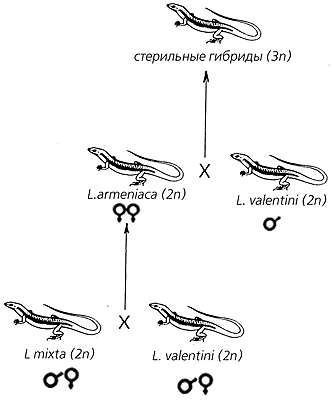
Схема гибридогенного образования (сетчатой эволюции) партеногенетического вида L.armeniaca
с последующим обратным скрещиванием и образованием триплоидных гибридных особей.К настоящему времени накопилась обширная литература, свидетельствующая о том, что именно в результате сетчатой эволюции образовались многие диплоидные и триплоидные виды не только различных ящериц, но также земноводных и рыб. Как явление, диаметрально противоположное процессу эволюционной дивергенции, любая гибридизация, казалось бы, не должна способствовать процессу видообразования, и всего лишь три десятилетия назад известный американский зоолог и эволюционист Э.Майр, основываясь на ряде теоретических предпосылок, полагал, что “гибридогенное видообразование хотя и часто постулировалось, но ни разу доказано не было”. Однако сегодня мы знаем уже десятки видов в различных группах животного и растительного мира, происхождение которых связано именно с процессом межвидовой гибридизации. Как здесь не вспомнить великого Гёте, утверждавшего устами своего Мефистофеля: “Теория, мой друг, мертва, но древо жизни зеленеет вечно”.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований. Грант №99-04-49334.
Литература
1 Darevsky I.S. // J. Ohio Herpetol. Soc. 1966. V.5. P.115—152.
2 Даревский И.С. Скальные ящерицы Кавказа. Л., 1967; Darevsky I.S., Kupriyanova L.A., Uzzell T. Parthenogenesis in reptiles // Biology of the Reptilia / Eds R.M.Dawly and J.P.Bogart. N.Y., 1985. V.15. P.412—526; Darevsky I.S. Herpetology: Current research of the biology of Amphibians and Reptiles // Proc. of the First World Congress of Herpetology / Ed. K.Adler. L., 1992. P.21—39; Даревский И.С. Эволюция и экология партеногенетического размножения у пресмыкающихся // Современные проблемы теории эволюции. М., 1993. С.89—109.
3 Астауров Б.Л. // Генетика. 1969. Т.7. С.129—149.
4 Uzzell T., Darevsky I.S. // Copeia. 1975. №2. Р.204—222.
5 Гречко В.В., Федорова Л.В., Федоров А.Н. и др. // Молекуляр. биология. 1997. Т.31. С.244—252; Гречко В.В., Рябинин Д.М., Федорова Л.В. и др. // Молекуляр. биология. 1998. Т.32. С.172—183; Grechko V.V., Ryabinin D.M., Fedorov A.N. et al. // Amphibia-Reptilia. 1977. V.18. P.407—418; Fedorov A.N., Fedorova L.V., Grechko V.V. et al. // J. Mol. Evol. 1999. V.48. P.69—76.
6 Ryabinina N.L., Grechko V.V., Darevsky I.S. // Genetica. 1998. V.34, P.1661—1667; Ryabinina N.L., Grechko V.V., Semenova S.K., Darevsky I.S. // Russ. J. Herpetology. 1999. V.6. P.55—60.
7 Murphy R., Darevsky I.S., MacCulloch R. et al. // Genetica. 1997. V.101. P.125—130.
 Август 2000 |