№9, 2001 г.
© И.В.ДовгальПростейшие — обитатели пограничного слоя
И.В.Довгаль
Игорь Васильевич Довгаль, к.б.н., ст.н.сотр. отдела фауны и систематики беспозвоночных животных
Института зоологии им. И.И.Шмальгаузена Национальной академии наук Украины.Рисунки Я.Шейбеля, А.В.Янковского.
На любом погруженном в воду объекте, как правило, формируются сообщества прикрепленных одноклеточных организмов, относящихся к царству простейших, или протистов (Protista), — многочисленной, но все еще недостаточно изученной группы беспозвоночных животных. Такие сообщества, обитающие на твердых субстратах в пресных водоемах, называют перифитоном, в морях — биообрастаниями, на поверхности водорослей и высших водных растений — эпифитоном, на водных животных — комменсальными сообществами, или эпибионтами, и т.д. По нашим оценкам, из примерно 100 тыс. известных видов протистов не менее 10% ведут сидячий образ жизни. Прикрепленные формы есть среди различных групп простейших: солнечников, фораминифер, радиолярий, гетеротрофных и автотрофных жгутиконосцев, инфузорий. И это неудивительно, ведь сидячий образ жизни дает организмам определенные преимущества — на обтекаемой водой поверхности они всегда обеспечены пищей.
Представители разных групп простейших, ведущие сидячий образ жизни. Среди них преобладают седиментаторы — организмы, заглатывающие взвешенные пищевые частицы (например, бактерий). Солнечники и суктории — в основном хищники: первые ловят и поедают добычу с помощью псевдоподий (аксоподий), а вторые — с помощью специализированных щупалец.Несмотря на распространенность, сообщества сидячих простейших до сих пор плохо изучены. Очень трудно выявить, какие именно факторы среды влияют на их развитие в естественных местообитаниях, нелегко воспроизвести условия подобных биотопов и в лаборатории. Поэтому обычно рассматривается весь комплекс воздействующих на сидячих простейших факторов (приливно-отливные и речные течения, конвекционные потоки и т.п.), хотя немало ученых считает, что основное значение среди них имеют гидродинамические нагрузки.Впервые серьезно этой проблемой занялись британские исследователи — специалист по гидродинамике Н.Р.Сильвестр и протозоолог М.А.Слей [1]. Авторы исходили из того, что прикрепленные водные организмы размером менее 1 мм находятся в пределах гидродинамического пограничного слоя, т.е. того слоя воды, в котором она затормаживается у обтекаемой ею поверхности. На основание прикрепленного объекта действует напряжение сдвига, сталкивающее его с поверхности субстрата. При этом величина интегральной силы (в отечественной литературе используется другой термин — гидродинамический упор) возрастает с увеличением диаметра объекта и в перпендикулярном направлении от поверхности субстрата, поэтому гидродинамические нагрузки наибольшее значение имеют для стебельчатых форм. Этим в основном ограничивались представления о гидродинамике сидячих простейших.
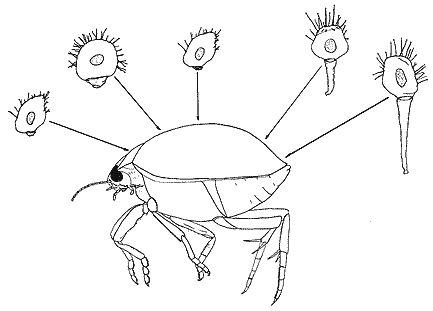
На разных участках верхней поверхности тела водного жука Hyphydrus ovatus сосущая инфузория Discophiya lichtensteinii имеет разную длину стебелька. Оказалось, что это связано с характеристиками гидродинамического пограничного слоя.Несколько лет назад автору этих строк удалось собрать около 30 экземпляров водяных жуков Hyphydrus ovatus из разных биотопов. Надкрылья жуков были густо покрыты инфузориями-сукториями Discophrya lichtensteinii. Даже визуально было заметно, что размер инфузории зависел от места ее расположения: чем ближе к заднему концу тела жука, тем длиннее был ее стебелек. Естественно было предположить, что такая закономерность связана с распределением гидродинамических нагрузок по поверхности надкрыльев. Необходимые расчеты помог выполнить сотрудник Института гидромеханики В.А.Кочин. Действительно, длина стебелька сукторий оказалась достоверно связана с параметрами пограничного слоя. Проанализировав опубликованные в научной литературе сведения об изменчивости размеров инфузорий в зависимости от их локализации, мы окончательно утвердились в своем предположении. И тогда нас стали интересовать не столько сами факторы, действующие на организмы в пограничном слое, сколько механизмы их адаптации к этим факторам [2]. Оказалось, что пограничный слой как биотоп имеет определенную структуру независимо от типа субстрата, что позволило рассматривать его в качестве комплекса потенциальных местообитаний, т.е. своеобразной адаптивной зоны сидячих простейших [3].
 |
 |
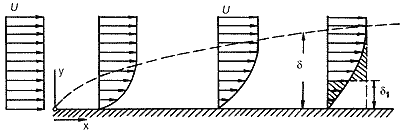
Электронные микрофотографии суктории Discophrya elongata на ноге водного клопа Ranatra linearis. У верхней поверхности ноги (слева) направление потока, видимо, относительно постоянно, поэтому инфузории наклонены в одну сторону. Увел. ґ600. У другой части той же ноги (справа), вероятно, возникают турбулентные вихри, соответственно здесь сукториям трудно “сориентироваться”, и они расположены хаотично. Увел. ґ200.Структура пограничного слоя как адаптивной зоныЧтобы не навлечь на себя гнев читателя-физика, сразу оговорюсь: употребление физического понятия “пограничный слой” в качестве адаптивной зоны в какой-то степени биологическая вольность. Как известно, существуют два подхода к определению размера пограничного слоя. В одних случаях принимается, что эта зона начинается там, где скорость течения на 1% (или 5%) меньше скорости потока окружающей жидкости, в других — на расстоянии от обтекаемой поверхности, на которое отклоняются линии тока жидкости (толщина вытеснения) [4]. Отдельно рассматривается так называемый диффузный пограничный слой. Все эти величины в нашем случае представляют интерес как составляющие пространственной структуры адаптивной зоны. Поэтому в дальнейшем речь пойдет о слоях, размер которых по нормали к поверхности совпадает с той или иной толщиной (диффузного пограничного слоя, толщиной вытеснения и т.п.), а не о величинах, принятых в гидродинамике.
Если пограничный слой ограничить толщиной вытеснения, то следует учесть, что в этих пределах практически отсутствует течение и гидродинамический упор ничтожен. Здесь же формируется диффузный пограничный слой, в пределах которого из-за отсутствия течения возможна только молекулярная диффузия, скорость которой во много раз ниже, чем у конвекционной диффузии. Таким образом, адаптивная зона включает слой, непосредственно соприкасающийся с обтекаемой поверхностью (для простейших — субстратом) и ограниченный толщиной вытеснения. Здесь из нагрузок действует лишь напряжение сдвига, кроме того, возможна только молекулярная диффузия. Далее, до края “классического” пограничного слоя (Сильвестр и Слей использовали “1%-ю” границу) скорость потока повышается, и начинает действовать и гидродинамический упор. Кроме того, в этом слое возможна конвекционная диффузия, что, конечно, существенно сказывается на транспорте питательных веществ. Такова вертикальная структура адаптивной зоны.
У поверхности обтекаемого тела (в данном случае пластины) в пограничном слое (d) скорость потока (U) постепенно снижается. Если в качестве размера пограничного слоя принимать толщину вытеснения (d1), то в ней значение скорости исчезающе мало — происходит так называемое “прилипание” жидкости.В направлении от переднего к заднему концу обтекаемого объекта толщина пограничного слоя увеличивается, а величина напряжения сдвига уменьшается. Если речь идет не об идеальном субстрате (пластине), а о реальном объекте, то ближе к заднему его концу происходит отрыв пограничного слоя, и здесь появляются турбулентные вихри, что создает дополнительную нагрузку для сидящих на поверхности организмов. Так формируется горизонтальная структура адаптивной зоны.Безусловно, помимо гидродинамических нагрузок в пограничном слое должны учитываться и традиционно рассматриваемые факторы — температура, концентрация кислорода, растворенного органического вещества и т.п. Для обитателей пограничного слоя имеют значение даже форма поверхности и другие свойства субстрата. Несомненно, характеристики пограничного слоя будут различны у поверхности камня на дне реки, возле таллома водоросли, растущей в приливно-отливной зоне, или у надкрыльев плывущего жука. В последнем же случае условия постоянно меняются, так как жук перемещается с разной скоростью и в разных направлениях, тем не менее это не мешает инфузориям не просто удерживаться на его поверхности, но и жить, питаясь и производя на свет себе подобных. Как же простейшие сумели освоить столь сложные для существования биотопы?
Адаптации простейших к жизни в пограничном слое
Представители разных групп простейших неоднократно и независимо переходили к сидячему образу жизни, но при этом сталкивались с одними и теми же проблемами. Первоначально подвижные временно прикрепляющиеся формы оказывались в пределах толщины вытеснения и подвергались воздействию только напряжения сдвига. Чтобы закрепиться на субстрате такие протисты должны были обладать органеллами секреции клейких веществ. Дальнейшая их эволюция проходила, очевидно, в направлении усиления функций таких органелл. Помимо этого, селективное преимущество, вероятно, получали формы с расширенным основанием клетки или с базальными ее выростами (для противодействия напряжению сдвига необходима большая площадь контакта с субстратом). В результате в разных группах сформировались распластанные по субстрату, иногда разветвленные, формы.
На этом этапе освоения адаптивной зоны прикрепившиеся организмы оказались в пределах диффузного пограничного слоя. Выход за его границу в область гораздо более быстрой конвекционной диффузии давал простейшим существенные преимущества в обеспеченности пищей. Вероятно, поэтому следующим этапом эволюции прикрепления у некоторых форм стало увеличение их тела. Так, у инфузорий появился стебелек (за счет усиления все той же секреции), позволивший им подняться над субстратом. Но при этом прикрепленные организмы начали подвергаться воздействию еще одной нагрузки — гидродинамического упора. Напомним, что его значение возрастает вместе с увеличением размеров обтекаемого тела, причем более всего в зоне соединения стебелька и клетки. Понятно, что необходимо было как-то обеспечить прочность именно этой зоны.
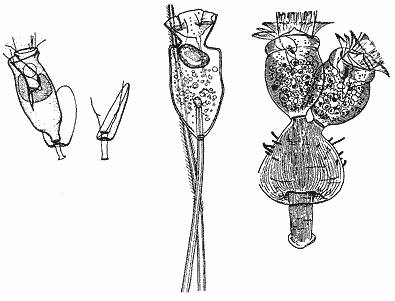
Существуют три возможных способа укрепить место соединения стебелька и клетки, и, что интересно, все они реализованы простейшими. Первый вариант — чашевидное расширение вершины стебелька, закрывающее основание клетки. Такое расширение известно у инфузорий (сукторий и перитрих) и называется полураковиной, назначение которой раньше убедительно никто не мог объяснить. В дальнейшем из полураковины сформировалась раковина, покрывающая клетку полностью [5]. Утолщение стебелька иной формы (физон) также известно у инфузорий и тоже выполняет функцию защиты зоны соединения. Второй вариант — внедрение верхней части стебелька внутрь клетки. Подобная структура (эндостиль) есть у сидячих инфузорий — хонотрих и сукторий. Третий возможный вариант — внедрение базальной части клетки внутрь стебелька. Он также реализован у инфузорий (перитрих) в виде проходящего по стебельку сократимого выроста клетки — спазмонемы. Первоначальной функцией спазмонемы, вероятно, было обеспечение прочности соединения стебелька и клетки, но в этом выросте оказались структуры, которые позволили стебельку сокращаться (микротрубочки и цистерны эндоплазматической сети).
Для сидячих простейших (особенно инфузорий) характерно разнообразие структур, защищающих соединение стебелька и клетки. Они развиты тем больше, чем в более экстремальных гидродинамических условиях обитает организм. У перитрихи Pyxidiella tectiformis, обитающей на жабрах ельца, стебелек снабжен асимметричной полураковиной ѕ скорее свидетельство адаптации к гидродинамическому упору, чем защита от хищников. Сходную функцию, вероятно, выполняет утолщение стебелька (физон) другой перитрихи Foisnerella physonica, обитающей на жабрах морской изоподы Gnorimosphaeroma noblei. У хонотрихи Oxychona multifida с рачков рода Nebalia проникающий внутрь клетки вырост стебелька (эндостиль) занимает почти треть клетки, обеспечивая высокую прочность соединения.Таким образом, у сидячих простейших (в первую очередь инфузорий) сформировалось удивительное разнообразие прикрепительных органелл, которые позволили им приспособиться к гидродинамическим нагрузкам.Известны и другие формы адаптации простейших к сидячему образу жизни — например, приспособление к особенностям структуры поверхности субстрата (это относится к видам, которые поселяются в сочленениях тела или конечностей хозяев-членистоногих) или усиление опорной и защитной функции покровов клетки (в частности, у инфузорий хонотрих и сукторий усиление функции кортекса привело к образованию кутикулы *, часто снабженной различными шипами и другими образованиями).
* Кутикула (от лат. cuticula — кожица) — плотный внеклеточный покров на поверхности эпителия многоклеточных животных. Для простейших принято использовать другой термин — пелликула, однако для сидячих инфузорий, которых мы рассматриваем, характерно усиление наружных структур, и у них формируется именно кутикула (а пелликула была еще у их предков и никуда не делась). Этот термин уже общепринят у протозоологов, и как раз о возникновении этой новой структуры у наших подопечных (которая есть еще и у кишечных инфузорий) идет речь [6]. Более того, полагают, что у автотрофных организмов переход к сидячему образу жизни привел к появлению у них клеточной оболочки, что в свою очередь считается одной из предпосылок возникновения многоклеточности у растений [7].
Наконец, вспомним и о собранных нами водяных жуках Hyphydrus ovatus, покрытых инфузориями Discophrya lichtensteinii, у которых длина стебелька зависела от их локализации на теле насекомого. Помимо тех гидродинамических нагрузок, которые анализировались британскими специалистами, эти инфузории испытывают дополнительные неблагоприятные нагрузки — турбулентность и нестационарность пограничного слоя. Чтобы справиться с нестационарностью, у них сформировался гибкий стебелек, позволяющий простейшему при возрастании гидродинамического упора отклоняться и прижиматься к субстрату. Приспособлением к турбулентности, по всей видимости, служит удлинение гибкого стебелька, что дает возможность двигаться вместе с обтекающей жидкостью, подобно тому, как длинные талломы морских водорослей изгибаются вместе с волнами прибоя.
С переходом к прикрепленному образу жизни связано и формирование специализированных органелл захвата и поглощения пищи простейших, но такие адаптации только косвенно связаны с гидродинамическими нагрузками.
Нетрудно заметить, что у простейших неподвижный образ жизни не связан, как у многоклеточных, с утратой каких-либо структур. Наоборот, разнообразие условий в пограничном слое вызвало развитие дополнительных органелл (в особенности прикрепительных). Более того, произошло усиление функции органелл питания, что привело к увеличению размеров клетки. Несомненно, все эти изменения имели прогрессивный характер [8]. * * *
Поселяясь в разных участках пограничного слоя и приспосабливаясь к таким условиям, протисты существенно снизили конкуренцию за субстрат и пищу. Например, разные виды сукторий, которые питаются свободноплавающими инфузориями, при поселении на неорганических субстратах образуют несколько групп по степени высоты над субстратом. Так они, вероятно, делят трофическую нишу. За счет различных адаптаций к напряжению сдвига простейшие по-разному распределяются и по поверхности субстрат. Таким образом, структура пограничного слоя определяет и пространственную структуру соответствующих сообществ.
Безусловно, это пока еще только схематическая картина условий обитания простейших на обтекаемой поверхности. Но уже первые попытки подойти к изучению сидячих протист с позиций гидродинамики позволили наполнить более конкретным содержанием формулировку “сидячий образ жизни простейших”.
Литература
1. Silvester N.R., Sleigh M.A. // Freshwater Biology. 1985. V.15. Р.433—448.
2. Довгаль И.В., Koчин В.А. // Вестн. зоологии. 1995. Т.29. №4. С.19—24.
3. Довгаль И.В., Koчин В.А. // Журн. общ. Биологии. 1997. Т.58. №2. С.67—74.
4. Шлихтинг Г. Теория пограничного слоя. М., 1974.
5. Довгаль И.В. // Вестн. зоологии. 1998. Т.32. №2. С.18—29.
6. Герасимова З.П.// Зоол. журн. 1989. Т.LXVIII. Вып.4. С.5—17.
7. Масюк Н.П. Эволюционные аспекты морфологии эукариотических водорослей. Киев, 1993.
8. Довгаль И.В. // Журн. общ. биологии. 2000. Т.61. №3. С.290—304.