№9, 2004 г.
№9, 2004 г.
№9, 2004 г.
© Попадьин К.Ю., Мамирова Л.А.
ИСТОРИЯ ОДИНОКОЙ ХРОМОСОМЫ
К.Ю. Попадьин, Л.А. Мамирова
Константин Юрьевич Попадьин, асп. каф. общей экологии биофака МГУ.
Лейла Абдумажетовна Мамирова, асп. каф. биохимии того же факультета.У большинства организмов с генетическим определением пола наряду с аутосомными хромосомами существуют половые, у млекопитающих - X и Y, а у птиц - W и Z. При этом у млекопитающих самки гомогаметны (ХХ), а самцы гетерогаметны (XY). Аналогичная ситуация наблюдается у мух Drosophila и некоторых растений. А вот у птиц картина обратная: самки гетерогаметны (WZ), a самцы гомогаметны (ZZ). Нас интересуют одинокие половые хромосомы, те, которые никогда не бывают вдвоем, поскольку определяют гетерогаметный пол: y млекопитающих это Y-хромосома, а y птиц - W. Здесь для простоты мы будем называть их Y-хромосомой, тем более что разговор пойдет в основном о млекопитающих.
Чем же замечательна одинокая хромосома? Дело в том, что все гомологичные хромосомы благодаря кроссинговеру, происходящему во время мейоза, обмениваются гомологичными участками друг с другом. А между половыми хромосомами X и Y, несущими поло-специфические гены самца и самки, генетического обмена не должно быть во избежание появления гермафродитов или неплодовитых особей среднего рода. Однако у самок X-хромосомы подвергаются кроссинговеру (поскольку их две), а вот Y-хромосома всегда одна!
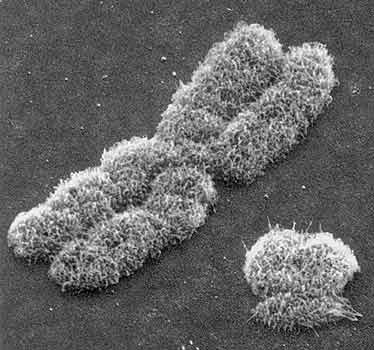
Электронная микрофотография X- и Y-хромосомы (справа) человека.
Увеличение 10 тыс. раз [1].Наличие или отсутствие кроссинговера у хромосом интересно в связи с эволюцией полового и бесполого размножения у всех организмов. Как известно, при бесполом размножении потомство производится путем деления (митоза) единственной клетки, т.е. каждая особь имеет лишь одного родителя, гены которого наследуются неизменно, за исключением вновь появившихся мутаций. При половом размножении благодаря кроссинговеру, независимому расхождению гомологичных хромосом и слиянию гамет при оплодотворении происходит рекомбинация генов, и потому генотип потомства отличается от родительского. Видимо, именно генетическое разнообразие потомства обеспечивает половому размножению (в отличие от бесполого) ряд эволюционных преимуществ [1]. Бесполых организмов очень мало (примерно 1% всех видов), да и те, как правило, представляют собой эволюционный тупик [2]. Отсутствие рекомбинации у Y-хромосомы позволяет рассматривать ее как "бесполый* организм", который по правилам эволюционной генетики должен деградировать. Вымрет или не вымрет наша Y-хромосома (здесь мы обращаемся ко всем самцам Homo sapiens, а также ко всем сочувствующим)? Этот важный вопрос мы и ставим основной целью данной работы.
* Пусть читатель простит нас за следующий каламбур - "бесполая" половая Y-хромосома, однако это именно так: несмотря на то, что Y-хромосома несет на себе гены, специфичные для самцов, сама по себе она нерекомбинирующая, т.е. "бесполая".* * * Итак, отсутствие кроссинговера приводит к постоянному и полному сцеплению всех генов вместе, в результате чего единицей эволюции становится целая хромосома, а не отдельный ген, как это наблюдается у хромосом, подвергающихся кроссинговеру (все аутосомы и X-хромосома). Такое простое утверждение налагает существенный отпечаток на эволюцию Y-хромосомы: скорость фиксации вредных мутаций увеличивается, а благоприятных уменьшается, что в конечном итоге может привести к исчезновению функциональных генов на Y-хромосоме [3]. Принято выделять несколько ключевых механизмов, ответственных за деградацию Y-хромосомы: храповик Меллера, фоновый отбор, эффект Хилла-Робертсона и эффект попутного транспорта. Накопление вредных мутаций на Y-хромосоме по большей части происходит за счет работы храповика Меллера [3], поэтому этот процесс стоит разобрать поподробнее.Случайное вымирание всех наилучших (без вредных мутаций) Y-хромосом в популяции человека возможно только в том случае, если численность этих хромосом невысока, и чем их меньше, тем быстрее они будут исчезать. Для приблизительной оценки численности самцов, способных к размножению и обладающих наилучшей Y-хромосомой, удобно использовать следующую формулу [4]:
no = Ne-u/s,
где:
no - искомая численность оптимальных Y-хромосом;Полагая, что N в популяциях млекопитающих часто имеет порядок нескольких сотен тысяч, u = 0.015 и s = 0.0015 [3], можно найти no:
N - эффективная численность всех Y-хромосом в популяции;
u - количество новых слабо вредных мутаций, появляющихся в Y-хромосоме в каждом поколении;
s - вредность мутации (коэффициент отбора), если s = 0, мутация нейтральная, если s = 1, мутация летальная.no = 500 000·e-0.015/0.0015 ~ 23. Таким образом, при данных условиях всего 23 самца обладают идеальной Y-хромосомой. Если по какой-либо причине они не оставят потомства, то в отсутствие рекомбинации восстановить случайно утраченную идеальную Y-хромосому будет невозможно (поскольку обратные мутации, изменяющие мутантный аллель в аллель дикого типа, крайне редки и их можно игнорировать). После этого оптимальной станет Y-хромосома с одной вредной мутацией, далее (после случайного исчезновения и этих хромосом) с двумя, тремя и так далее. Таким образом, благодаря храповику Меллера постепенно будут накапливаться вредные мутации. Как быстро может работать этот механизм? Из этой математической модели при рассмoтренных параметрах следует, что очередная вредная мутация фиксируется в среднем один раз за 750 поколений [3]. Однако чем больше эффективная численность популяции и чем меньше отношение u/s, тем больше численность оптимальных хромосом и тем маловероятнее их исчезновение.
В подтверждение работы храповика и/или остальных трех процессов у большинства видов Y-хромосомы становятся более гетерохроматиновыми и генетически инертными: количество функциональных генов у них быстро уменьшается. X- и Y-хромосомы млекопитающих произошли от аутосомной пары гомологичных хромосом не ранее чем 300 млн лет назад, т.е. после того, как птицы уже отделились от ствола, ведущего к млекопитающим, и не позднее чем 170 млн лет назад, когда уже появились самые древние яйцекладущие млекопитающие [5]. Полагая, что количество генов на древних Y- и X-хромосомах было одинаково и оно осталось неизменным в X-хромосоме (сегодня в ней около 1500 генов), а число разных функциональных генов Y-хромосомы постоянно уменьшалось и сейчас меньше 30 [6], можно подсчитать темп деградации Y-хромосомы [7]. Почти за 300 млн лет Y-хромосома потеряла около 1470 генов, т.е. примерно 5 генов за каждый миллион лет. При данном темпе деградации оставшиеся гены должны исчезнуть примерно за 6 млн лет.
Деградируют не только Y-хромосомы млекопитающих - совершенно аналогичная ситуация наблюдается на Y-хромосомах растений [8] и дрозофилы. Особенно поучителен пример с нео-Y-хромосомой Drosophila miranda [9]. Она образовалась примерно 2 млн лет назад в результате слияния аутосомы с Y-хромосомой. В результате такой перестройки эти "аутосомные" гены стали наследоваться вместе с генами Y-хромосомы, т.е. "бесполо", и нео-Y-хромосома начала терять функциональные гены. Независимая деградация Y-хромосом разных организмов дополнительно подтверждает универсальность недостатков "бесполого" размножения и заставляет задуматься над будущим половой хромосомы человека.
Однако темп деградации Y-хромосомы непостоянен. Вначале его скорость высока, но постепенно, по мере уменьшения самой Y-хромосомы, она замедляется, поскольку сокращается количество функциональных генов; следовательно, снижается темп мутирования (u), что увеличивает численность оптимальных хромосом в популяции (см. формулу). Поэтому вышеприведенные расчеты о полном вымирании Y-хромосомы человека через 6 млн лет, скорее всего, не заслуживают внимания, поскольку современный темп деградации Y-хромосомы должен быть сильно замедлен (но не равен нулю).
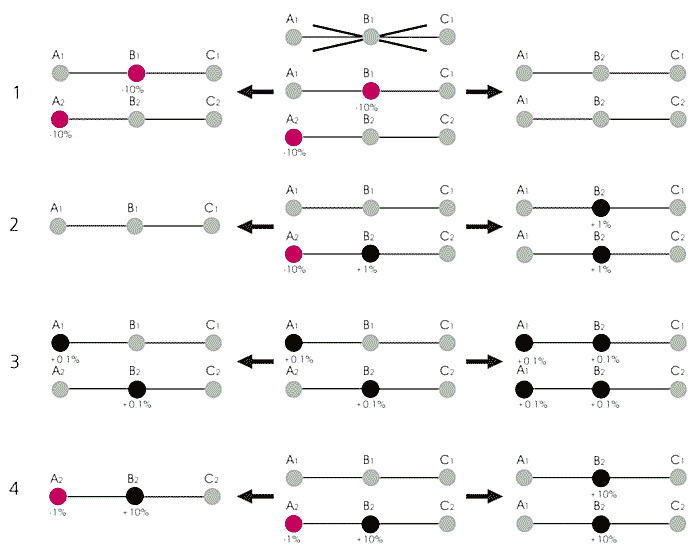
Схема, иллюстрирующая недостатки нерекомбинирующей хромосомы [3, 12, с изменениями]. A, B, C - разные гены. В центре - возможные комбинации новых мутаций, справа - в рекомбинирующих хромосомах, слева - в Y-хромосоме. Проценты показывают коэффициент отбора новых аллелей: + благоприятная мутация (красный цвет), - вредная мутация (черный цвет). Нейтральные аллели отмечены серым цветом.1. Храповик Меллера - необратимое накопление слабо вредных мутаций в результате случайной потери из популяции всех наилучших (с минимальным количеством вредных мутаций) Y-хромосом. Отмечена невозможность воссоздания утраченной хорошей хромосомы в отсутствие кроссинговера.
2. Фоновый отбор - нейтральная или слабо благоприятная мутация может зафиксироваться в популяции только в отсутствие в данной хромосоме вредных мутаций. В противном случае любая новая мутация будет элиминирована вместе с уже существующей вредной.
3. Эффект Хилла-Робертсона - замедление элиминации слабо вредных и фиксации слабо благоприятных мутаций из-за связи отбираемых аллелей друг с другом. Эффект работает только при малых значениях коэффициента отбора, когда действие генетического дрейфа и обратных мутаций велико. Чем больше сцепленных сайтов, тем меньше приспособленность нерекомбинирующего генома. Соединение слабо благоприятных мутаций невозможно, а поскольку приспособленность двух типов организмов одинакова, частота этих аллелей будет зависеть от генетического дрейфа и обратных мутаций.
4. Эффект попутного транспорта - совместная эволюция благоприятных, нейтральных и вредных мутаций. Вредные и нейтральные мутации могут распространиться в популяции за счет сцепления их с благоприятной. Один из эффектов недавнего распространения благоприятной мутации в популяции бесполых организмов - резкое уменьшение изменчивости по нейтральным сайтам - аллели А1, В1 и С1 потеряны.
В недавних работах обнаружены новые интересные особенности Y-хромосомы человека и шимпанзе, подающие дополнительную надежду всем ее обладателям [6, 10, 11]. Они открывают второй механизм адаптации, позволяющий даже в отсутствие кроссинговера эффективно бороться против накопления слабо вредных мутаций.
Авторы этих исследований делят Y-хромосому на три участка.
Первый (X-transposed) мигрировал с X-хромосомы на Y-хромосому в последние несколько миллионов лет. До сих пор он весьма похож на гомологичный участок X-хромосомы и содержит всего два гена.
Второй (X-degenerate) участок более древнего происхождения и представляет собой реликт древней аутосомы, от которой произошли половые хромосомы. 16 белок-кодирующих генов, содержащихся в нем, как правило, широко экспрессируются в разных тканях организма, продуцируя изоформы, немного отличающиеся от тех, что кодируются на X-хромосоме. Лишь один SRY-ген (sex-determining region of the Y), отвечающий за запуск каскада специфических для самцов генов, экспрессируется только в семенниках. В связи с большим возрастом данный участок служит моделью для изучения эволюции нерекомбинирующего генома, и на нем виден результат работы упомянутых выше четырех процессов (храповика Меллера, фонового отбора, эффектов Хилла-Робертсона и попутного транспорта). Видимо, на протяжении 300 млн лет Y-хромосома уменьшалась за счет этого участка. Возможно, он деградирует до сих пор, но с сильно замедленным темпом.
Третий участок (Y-specific repetitive sequences), наибольший по размеру, состоит из множественных повторов каждого из девяти присутствующих там генов, большинство которых собраны в палиндромы* (последовательности, читающиеся от 5'-конца одинаково на обеих комплементарных цепочках ДНК). Данные гены Y-специфические, поскольку давно мигрировали из различных аутосом и X-хромосомы, и большинство из них исчезло из первоначального местоположения. После миграции в Y-хромосому все они амплифицировались. Функции этих генов непосредственно связаны со сперматогенезом, поэтому экспрессируются они преимущественно в семенниках.
* Пример короткого палиндрома 5'GCCGGC3' - 3'CGGCCG5' . Длинные палиндромы, включающие в себя целые гены, образуются в результате дупликации и последующей инверсии одной из дуплицированных последовательностей.Для генов третьего участка характерен медленный темп молекулярной эволюции. С момента дивергенции человека и шимпанзе, произошедшей примерно 5 млн лет назад, они разошлись лишь на 1.4%, да и те обусловлены в основном нейтральными мутациями (синонимичные замещения). Более того, повторы каждого гена внутри палиндрома идентичны друг другу более чем на 99.9%! Небольшое расхождение, наблюдаемое между человеком и шимпанзе, а также между всеми повторами каждого гена, нельзя объяснить интенсивным отбором, поскольку даже нейтральные Alu последовательности внутри палиндромов показывают очень низкий темп эволюции.
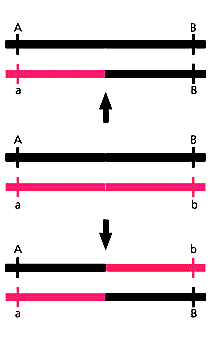
Схема гомологичной рекомбинации: в центре - две гомологичные последовательности ДНК, внизу - кроссинговер, или обмен, вверху - генная конверсия, или замещение (B заместил b). Оба процесса происходят во время мейоза и реже при митозе.Скорее всего, повторы каждого гена достаточно часто копируются с какого-то определенного шаблона, в результате чего независимое накопление мутаций в них прекращается. Такой процесс называется генной конверсией между копиями генов внутри Y-хромосомы.
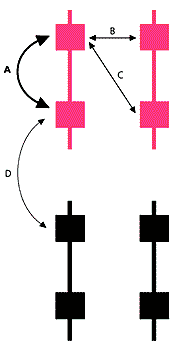
Генная конверсия - это гомологичная рекомбинация, при которой, в отличие от обмена участками при кроссинговере, один участок ДНК замещается другим. Наиболее подробно такой тип рекомбинации изучали на грибах, где впервые показали замещение генов между гомологичными хромосомами и реже между сестринскими хроматидами во время мейоза. Позже появились сведения о том, что генная конверсия возможна между дуплицированными копиями гена на одной хромосоме (как в Y-хромосоме), а также во время митоза.
Почему все Y-специфичные гены амплифицированы и каков эволюционный смысл интенсивной генной конверсии между копиями одного и того же гена? Имеются два не исключающих друг друга объяснения.
Первое состоит в том, что амплификация данных генов необходима для интенсивной экспрессии в семенниках. Множественные копии в отсутствие генной конверсии будут независимо накапливать мутации и дивергировать. Накопление вредных мутаций и снижение функциональной активности некоторых копий будет компенсироваться нормальными генами. В результате этого процесса количество функциональных копий снова уменьшится и станет недостаточным для необходимого уровня экспрессии. Генная конверсия между копиями гена позволит сохранять его в консервативном состоянии. При появлении новой мутации в одной из копий в результате конверсии мутация либо вытеснится оригинальной последовательностью, либо, напротив, размножится. В первом случае новая мутация просто исчезнет, как будто она заместилась обратной мутацией или подверглась репарации. Во втором же - увеличение числа копий мутантного повтора усилит его вредный эффект (s), который к тому же не будет компенсироваться нормальными копиями данного гена. Значит, такая Y-хромосома станет менее функциональной, и сперматозоид с большой вероятностью элиминируется. Таким образом, мутация все равно исчезнет из популяции, но на этот раз за счет отсутствия потомства у данного индивидуума. Важно отметить, что увеличение общего вреда (s) при конверсии в сторону мутантной последовательности, равно как и уменьшение темпа мутирования (u) при конверсии в сторону оригинальной последовательности замедлят работу храповика (см. формулу).
Второе объяснение связано с существованием преимущественно однонаправленной генной конверсии. Выше мы предполагали наличие случайной конверсии, когда вероятность замещения одной последовательности другой равна вероятности обратного процесса. Однако, как правило, генная конверсия преимущественно однонаправленная, т.е. вероятность прямого замещения не равна вероятности обратного. Применительно к Y-хромосоме это может означать, что у мутантной последовательности шансы на амплификацию много меньше, нежели у оригинальной. Значит, возможно существование дополнительного механизма, уменьшающего распространение новой (и вредной, и благоприятной) мутации. Хотя наличие преимущественно однонаправленной генной конверсии в геномах многих организмов - факт хорошо доказанный [12], механизмы и эволюционный смысл этого процесса еще обсуждаются. Одно из объяснений касается элиминации новых мутаций: поскольку большинство их смещено в сторону аденина и тимина, то генная конверсия, сдвинутая в сторону гуанина и цитозина, будет автоматически элиминировать новые мутации [12]. При таком сценарии присутствие нескольких копий каждого гена диктуется необходимостью ферментов генной конверсии "сравнивать" нуклеотиды разных копий.
Как мы уже говорили, оба объяснения не исключают друг друга, и однонаправленная генная конверсия будет лишь уменьшать частоту появления мутантов с низкой приспособленностью (после конверсии в сторону мутантного аллеля).
Медленный темп эволюции генов третьего участка (небольшая скорость фиксации новых мутаций) подтверждается заниженным временем дивергенции в филогенетическом древе, восстановленном по гаплотипам Y-хромосом [6,13]. Также большую изменчивость репродуктивного успеха самцов можно отчасти объяснить именно мутациями в ключевых генах Y-хромосомы, при этом пониженная плодовитость и приведет к исчезновению данного мутантного гаплотипа Y-хромосомы [13].
На первый взгляд, низкий темп эволюции генов Y-хромосомы противоречит широко принятому мнению о быстрой эволюции генов, связанных с половым отбором [14]. Убедительно доказана большая скорость эволюции репродуктивных генов самцов у предков людей [15], а у дрозофилы обнаружено быстрое распространение нового благоприятного репродуктивного гена самцов [16]. Таким образом, утверждение о максимальной скорости положительного (дарвиновского) отбора в генах, отвечающих за репродуктивные свойства самцов, хорошо обосновано и с теоретической [14], и с практической [15-17] сторон.
Как же это согласуется с нашими рассуждениями о Y-хромосоме? Дело в том, что далеко не все гены, отвечающие за репродуктивные свойства самцов, находятся на Y-хромосоме: большинство их "разбросано" по всем аутосомам, и достаточно много в X-хромосоме [18, 19]. На Y-хромосоме должен присутствовать лишь основной ген, делающий самцов самцами, - SRY-ген. Именно он запускает индукцию всех остальных генов, специфичных для самцов. Значит, высокая скорость положительного отбора обусловлена не Y-хромосомными генами, а аутосомными и X-хромосомными, тогда как Y-специфичные гены (из третьего участка) как бы заморожены, и нам не известно ни одного убедительного примера селективного выметания, связанного с распространением благоприятной мутации на этих генах. Поскольку любая благоприятная мутация - событие уникальное, говорить об отсутствии положительного отбора на Y-cпецифичных генах из-за генной конверсии нельзя. Корректнее утверждать, что высокий темп эволюции признаков, сцепленных с полом, обусловлен аутосомными и X-хромосомными генами.
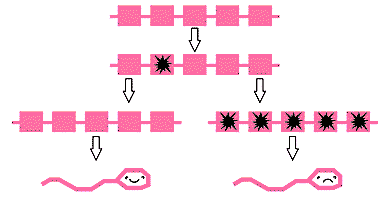
Гипотетическая схема генной конверсии между копиями одного и того же гена. Возможно, что вновь появившаяся в одной из пяти копий (цветная звездочка) вредная мутация в результате случайной генной конверсии сразу же "репарируется" (слева). Если же мутантная копия амплифицируется (это может произойти в одном из пяти случаев при случайной генной конверсии и намного реже при сдвинутой генной конверсии), вредный эффект усилится, что приведет к пониженной плодовитости ее носителя и, следовательно, к элиминации данной мутации (справа).Итак, история одинокой хромосомы показывает, что эволюция генов в отсутствие генетической рекомбинации действительно сопряжена с накоплением вредных мутаций и низкой скоростью распространения благоприятных. Однако, несмотря на это, Y-хромосома млекопитающих несет на себе функциональные гены и не уменьшается так быстро. Скорость эволюции (20 генов) второго участка замедлена просто в связи с малым количеством сцепленных генов (малый u), а все Y-специфичные амплифицированные гены (третий участок) имеют дополнительный адаптивный механизм в виде интенсивной генной конверсии (малый u и большой s). Поскольку эволюция генов Y-хромосомы согласуется с математической моделью храповика, можно заключить, что накопление вредных мутаций "бесполыми" генами - основной недостаток бесполого размножения у всех организмов.
Авторы выражают благодарность Дмитрию Филатову за рациональную критику
и Европейскому научному фонду и научной партеногенетической группе за приглашение на семинары, посвященные изучению партеногенеза.Работа поддержана Российским фондом фундаментальных исследований. Проект № 04-04-49623.
Литература
1. Попадьин К.Ю. // Журнал общей биологии. 2003. T.64. C.463-478.
2. Maynard Smith J. The evolution of sex. Cambridge, 1978.
3. Charlesworth B., Charlesworth D. // Phil. Trans. R. Soc. Lond. B. 2000. V.355. P.1563-1572.
4. Haigh J. // Theor. Pop. Bio. 1978. V.14. P.251-267.
5. Graves J.A.M. // TIG. 2002. V.18. P.259-264.
6. Skaletsky et al. // Nature. 2003. V.423. P.825-837.
7. Aitken R.J., Graves J.A.M. // Nature. 2002. V.415. P.963.
8. Filatov D., Moneger F., Negrutiu I., Charlesworth D. // Nature. 2000. V.404. P.388-390.
9. Steinemann M., Steinemann S. // Genetica. 2000. V.109. P.105-111.
10. Rozen et al. // Nature. 2003. V.423. P.873-876.
11. Willard H.F. // Nature. 2003. V.423. P.810-813.
12. Marais G. // TIG. 2003. V.19. P.330-338.
13. Cummins J. // Reprod. Fertil. Dev. 2001. V.13. P.533-542.
14. Gavrilets S. // Nature. 2000. V.403. P.886-889.
15. Wyckoff G.J., Wang W., Wu C. // Nature. 2000. V.403. P.304-308.
16. Nurminsky D.I., Nurminskaya M.V., De Aguiar D., Hartl D.L. // Nature. 1998. V.396. P.572-575.
17. Wang P.J., McCarrey J.R., Yang F., Page D.C. // Nature Genet. 2001. V.27. P.422-426.
18. Hurst L.D. // Nature. 2001. V.411. P.149-150.
19. Li W.-H. Molecular evolution. Sunderland, 1997.