№9, 2004 г.
№9, 2004 г.
№9, 2004 г.
© С.В. Наугольных
Преуспевающий реликт
С.В. Наугольных
Сергей Владимирович Наугольных, д. г.-м. н., вед. н. с. Геологического института РАН.
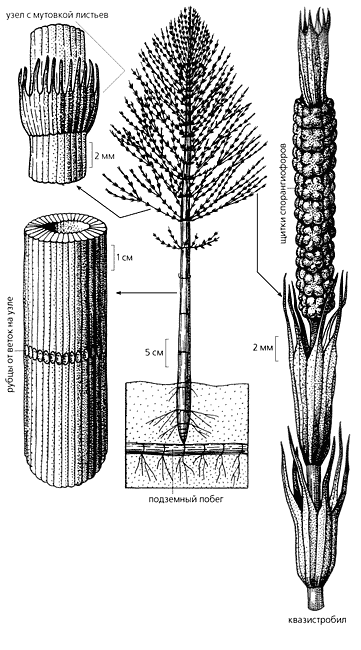
Ранней весной даже горожанин, наверное, замечал на обрывах канав, что проложены вдоль дорог, красновато-белые столбики, устремленные вверх. Это генеративные побеги хвоща полевого (Equisetum arvense). На конце побега находится стробил (с лат. - шишка) - орган размножения, в котором образуются споры. После их созревания надземная часть отмирает и появляется невысокое, 15-30-сантиметровое растение, похожее на маленькую елочку. Его ажурные боковые ветви, собранные мутовками, отходят от узлов рифленого полого стебля. Листья у хвощей превратились в мелкие чешуйки, прижатые к узлам и не способные к фотосинтезу; эту функцию выполняют зеленые стебли и ветви. Другие виды этих многолетних трав с жесткими, пропитанными кремнеземом стеблями, могут достигать метровой высоты, а у некоторых полегающих видов стебель иногда тянется на 9 м.
Род Equisetum (кстати, его латинское название происходит от двух слов: equis - лошадь, и seta, или saeta, - щетина, грива) насчитывает около 30 видов и распространен повсюду, за исключением Австралии и Новой Зеландии.
У человека, сидящего на берегу подтопленного заболоченного лесного озерца или речушки и смотрящего на густые прибрежные заросли хвощей с длинными трескучими членистыми стеблями, легко может возникнуть иллюзия, будто он находится в лесу, росшем в конце палеозойской эры, в пермском периоде. Близких и дальних родственников хвощей тогда было видимо-невидимо. Они так же, как их нынешние потомки, предпочитали селиться в хорошо увлажненных местах и, судя по известным местонахождениям растительных остатков этого возраста, нередко образовывали моновидовые сообщества.
Воистину палеозойская эра была периодом расцвета членистостебельных. Но из огромного разнообразия этих споровых высших растений до наших дней дожил всего лишь один род хвощей. Известны ли их предки, что они собой представляли и благодаря чему сумел сохраниться и столь преуспеть этот единственный род реликтовых растений? Об этом и пойдет речь.
Два кандидата
Почти в любом учебнике ботаники, да и в популярной литературе тоже, говорится, что хвощи ведут свою родословную от каламитов - гигантских споровых растений с членистыми стволами, достигавшими 20 м в высоту, а то и более. Сейчас каламитов относят к самостоятельному семейству Calamitaceae (или Calamostachyaceaе), которое обособляют в особый порядок Calamitales, называемый также Calamostachyales [1]. С каламитами современных хвощей сближает своеобразная ребристость ствола: против каждого ребра на междоузлии находится впадинка в узле, т.е. ребра на этих частях растения сдвинуты относительно друг друга. У других палеозойских членистостебельных ребра проходят через узлы побега прямо, без смещения. К таким растениям относятся, например, астеро-, или археокаламиты, клинолистники-сфенофиллы, а также представители семейства черновиевых, из которых наиболее известен формальный род паракаламитес (Paracalamites). Из перечисленных групп сфенофиллы - очень своеобразная группа споровых растений - особенно далеки от хвощей и потому не могут рассматриваться в качестве кандидатов на роль их филогенетических предшественников. Зато семейство черновиевых наравне с каламитами может претендовать на роль предков современных хвощей, несмотря на отличие в ребристости побега.
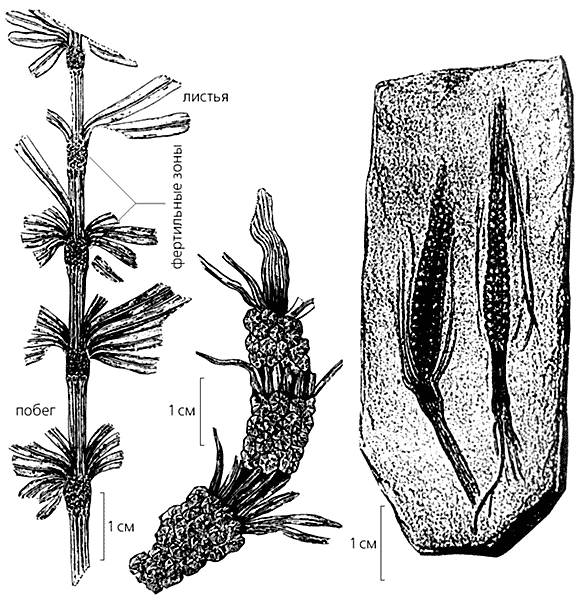
 Вегетативные и репродуктивные органы двух видов пермских черновиевых из Приуралья - филлотеки и паракаламитины. Слева направо: вегетативный побег филлотеки (Phyllotheca stenophylloides) в породе; реконструкция части вегетативного побега; репродуктивный орган эквизетиностахис с многоярусным расположением фертильных зон (рис. по фотографии [9]) и прорисовка разных морфологических частей паракаламитины (Paracalamitina striata). В последнем рисунке масштаб 2 мм.
Вегетативные и репродуктивные органы двух видов пермских черновиевых из Приуралья - филлотеки и паракаламитины. Слева направо: вегетативный побег филлотеки (Phyllotheca stenophylloides) в породе; реконструкция части вегетативного побега; репродуктивный орган эквизетиностахис с многоярусным расположением фертильных зон (рис. по фотографии [9]) и прорисовка разных морфологических частей паракаламитины (Paracalamitina striata). В последнем рисунке масштаб 2 мм.Черновиевые (Tchernoviaceae), составляющие самостоятельное семейство в порядке хвощевидных (Equisetales), произрастали в основном в каменноугольное и пермское времена на территории Ангариды, материка, который располагался в пределах современной северо-восточной Азии. Судя по многим признакам, черновиевые произошли от астерокаламитовых, живших в раннем карбоне [1].
Какие же палеозойские растения стояли в основании филогенетической цепочки, ведущей к современным хвощам? Каламиты или черновиевые?
Чтобы ответить на этот вопрос, надо понять, какие из предполагаемых предков наиболее сходны строением с хвощами.
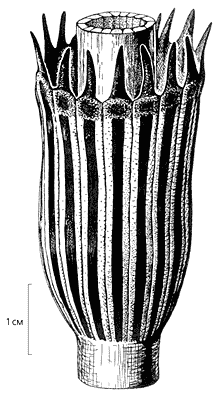
Спороношения. Известно, что весьма важным таксономическим признаком служит строение репродуктивных органов (у членистостебельных их называют спороношениями).
У каламитов эти органы образуют компактные собрания (стробилы) и внешне несколько напоминают стробилы современных хвощей. Однако если присмотреться к некоторым важным деталям в строении тех и других растений, выяснится, что между ними существует много различий. Главное из них заключается в том, что спорангиофоры хвощей расположены на фертильной оси один за другим и примыкают друг к другу, а у каламитов находятся в пазухах видоизмененных чешуевидных листьев - брактей.
Спороношения черновиевых во многом устроены иначе. У растений рода Equisetinostachys, одного из наиболее типичных в семействе черновиевых, спороношения больше, чем у каламитов, сходны со стробилами современного хвоща. Спорангиофоры закрыты щитками, края которых на незрелых генеративных органах плотно сомкнуты, а по мере созревания спорангиев загибаются кверху. Благодаря этому споры высыпаются, выходят наружу и разносятся водой и ветром.
У Equisetinostachys существовали и своеобразные черты строения спорошений. На верхушке побега располагался не единственный стробил, как у хвоща, а несколько фертильных зон образовывали сложную многоярусную конструкцию, разделенную мутовками стерильных листьев. Пучок листьев венчал и вершину фертильного побега эквизетиностахиса. Из этого следует, что верхушечные меристемы побега (в отличие от стробилов современного хвоща) были активными. Таким образом, спороношения Equisetinostachys не образовывали настоящих специализированных стробилов, а располагались непосредственно на побеге растения.
Формы роста. На реконструкциях палеозойских ландшафтов древние членистостебельные нередко изображаются как увеличенные во много раз копии современных хвощей. Однако такое представление ошибочно. Конечно, и у современных хвощей, и у их ископаемых родственников во внешнем виде много общего. Прежде всего, у них одинаковый - моноподиальный - тип ветвления, а ветви и листья собраны в мутовки. Но, по большому счету, тем сходство и заканчивается.
Формы роста каламитов еще в 20-х годах прошлого столетия реконструированы знаменитым немецким палеоботаником М.Хирмером [2]. (Его реконструкции вошли во многие учебники палеонтологии и ботаники.) По особенностям расположения ветвей на побеге Хирмер выделил несколько подродов каламитов (иногда их рассматривают в качестве самостоятельных родов): Stylocalamites, Crucicalamites, Diplocalamites, Calamitina. Позднее были добавлены еще несколько реконструкций, основанных на строении найденных стволов с ветвями и даже спороношениями, а также на результатах сравнительно-морфологических исследований [3].
 |
 |
 |
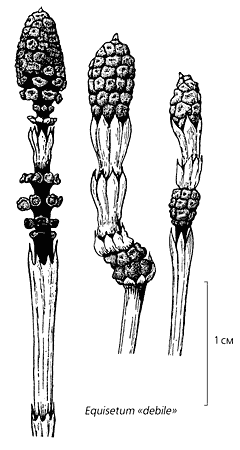
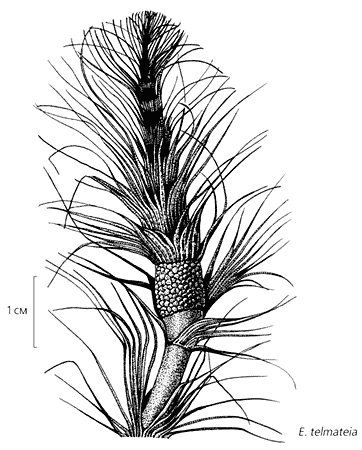
Форма аберрантных побегов у трех видов современного хвоща. У Equisetum «debile» это - фертильные зоны, такие же как у ископаемого эквизетиностахиса [10, с изменениями]. Вершина стробила у E.telmateia остается активной, и по существу он представляет собой фертильную зону, расположенную на междоузлии [11, с изменениями]. Стробилы E.hyemale располагаются на вершинах осевого и боковых побегов, чем напоминают растения двух мезозойских родов Equisetites и Neocalamites.
Относительно ангарских черновиевых было в общем-то понятно, что по меньшей мере некоторые из них представляли собой древовидные растения, поскольку вместе с побегами, листьями и спороношениями часто встречались фрагменты стволов довольно основательного диаметра - до 10-15 см. Как выглядели при жизни эти членистостебельные Ангариды, долгое время оставалось загадкой. Одна удачная находка помогла ее прояснить.
В представительном разрезе пермских отложений, расположенном по берегам (в основном, на левобережье) р.Кожим в Печорском Приуралье, мне посчастливилось найти крупный членистый ствол, явно относившийся к черновиевым. К узлам ствола прикреплялись облиственные побеги, на концах некоторых из них сохранились спороношения. После нескольких часов работы молотком и зубилом более полуметра ствола оказались свободными от скальной породы. По соотношению его диаметра и скорости уменьшения толщины удалось вычислить высоту растения - она должна была превышать 3.5 м. Эта находка послужила основой для первой графической реконструкции целого растения, названного Paracalamitina striata, из семейства черновиевых [4]. Прежде существовавшие реконструкции касались отдельных частей побегов [5] и спороношений [6] некоторых других представителей черновиевых.
Спороношения у Paracalamitina striata, как и у эквизетиностахиса, сохраняли активность терминальных меристем: на верхушках фертильных зон, которые можно условно назвать «квазистробилами», располагался все тот же хохолок из стерильных листьев. Но количество фертильных зон по сравнению с Equisetinostachys сократилось до одной, в очень редких случаях - двух. Другое пермское членистостебельное с намечающейся редукцией фертильных зон, расположенных на боковых побегах, было описано в 1983 г. С.В.Мейеном и Л.В.Меньшиковой как Sendersonia matura из верхнепермских отложений Кузбасса.
Древние признаки у современных хвощей
Один из эффективных способов корректировки филогенетических гипотез, высказываемых палеонтологами, - изучение изменчивости и аберраций современных организмов. Такие отклонения от нормы иногда повторяют черты строения отдаленных предков, потому что некоторые древние гены сохраняются в генотипе потомков в латентном, «спящем» состоянии, а затем, нередко под воздействием неблагоприятных факторов среды, могут активизироваться. Поэтому обнаружить в природе аберрантную форму - большая удача для палеоботаника.
Летом 1998 г. в приустьевой части Монастырского оврага (Ульяновская обл., правый берег Волги в 10 км выше по течению от г.Тетюши, близ с. Монастырское) мне попались на глаза несколько экземпляров современного хвоща зимующего (Equisetum hyemale), побеги которого существенно отличались от нормальных: стробилы располагались не только на главном осевом побеге, но и на двух-трех коротких боковых ветвях, идущих от самого верхнего узла. Ниже у всех экземпляров боковые ветви на стебле отсутствовали, что весьма характерно для этого вида хвощей. Стробил на основном побеге, как правило, был значительно лучше развит, чем на боковых ветвях. Высота собранных растений колебалась от 50 до 80 см при ширине стеблей 4 мм, а длина междоузлий варьировала от 38 до 65 мм.
Развитие стробилов на боковых побегах, для современных хвощей абсолютно нетипичное, свойственно мезозойским представителям родов Equisetites и Neocalamites. Примечательно, что у Neocalamites aff. сarrerei, описанного в 1958 г. В.П.Владимирович из угленосных триасовых отложений Челябинского бассейна, стробилы сидели на боковых ветвях по одному, как у Equisetum hyemale. У Equisetites arenaceous, еще одного мезозойского вида, стробилы тоже располагались на укороченных боковых побегах, но не по одному, а по три [7, 8].
Современные Equisetum имеют и другие аберрации. Известны экземпляры, генеративные органы которых разделены на несколько фертильных зон, фактически идентичных спороношениям уже упомянутого пермского черновиевого из рода Equisetinostachys. Иногда верхняя фертильная зона несет пучок стерильных листьев, аналогичных терминальному хохолку эквизетиностахиса.
У крупного современного хвоща Equisetum telmateia описаны экземпляры, у которых побег развивается из верхушки стробила. В этом случае последний по существу представляет собой фертильную зону, расположенную на междоузлии. У того же вида стробилы образуются и на боковых побегах, как у позднепермской Sendersonia matura, причем иногда верхушку венчает пучок стерильных листьев.
Примеров проявления древних признаков у современных хвощей в литературе можно найти множество, хотя в природе подобные отклонения встречаются редко.
Близкое родство
Ныне здравствующие хвощи, эти споровые растения из прошлых времен, случается, несут в себе, как уже сказано, необычные морфологические черты, которые были присущи предшественникам. Так, аберрация хвоща зимующего из Монастырского оврага повторяет форму репродуктивных органов триасовых хвощей. Фертильные зоны растений из пермских родов Equisetinostachys или Paracalamitina почти ничем не отличаются от тех аберрантных спороношений, которые образуются у современного хвоща, если терминальная меристема уже сформировавшегося стробила остается активной. В результате из верхушки генеративного органа вырастает побег, что совершенно несвойственно растению в норме. Такие аберрантные спороношения закладываются у современного Equisetum, как правило, на боковых ветвях, а не на основном побеге, как нормальные стробилы. И по этому признаку нынешние отклоняющиеся от обычной формы хвощи идентичны пермским членистостебельным семейства Tchernoviaceae (роды Equisetinostachys, Paracalamitina, Sendersonia). Следовательно, между семейством Equisetaceae и семейством позднепалеозойских ангарских черновиевых (Tchernoviaceae) существует прямая филогенетическая связь. По существу рекапитуляция примитивных генов, находящихся в латентном состоянии, превращает современные хвощи с аберрациями репродуктивных органов в уменьшенные во много раз копии их предков из палеозойской эры.
Если выстроить все формы, которые здесь обсуждались, в один ряд от древних видов к молодым, можно выявить неизменную морфогенетическую тенденцию - редукцию и генеративных, и вегетативных органов. Растения уменьшаются в размерах: от трех-четырехметровых паракаламитин до современных хвощей, большинство которых не превышает нескольких десятков сантиметров в высоту. Многочисленные фертильные зоны древних видов редуцируются до единственной, ее терминальная меристема теряет активность и в результате репродуктивные органы сохраняются только на верхушке побега - фертильная зона превращается в стробил. По всей видимости, редукция сопровождалась ускорением генеративных процессов: созревания репродуктивных органов и начала генеративной активности (фертильности) растения в целом. Это давало предкам хвощей преимущества в конкурентной борьбе и освоении подходящих экологических ниш.
Очень сходные тенденции известны и во многих других группах относительно примитивных высших растений: у папоротников, плауновидных, птеридоспермов, цикадовых и гинкговых. Не исключено, что редукция и олигомеризация наиболее значимы в эволюции споровых и голосеменных растений.
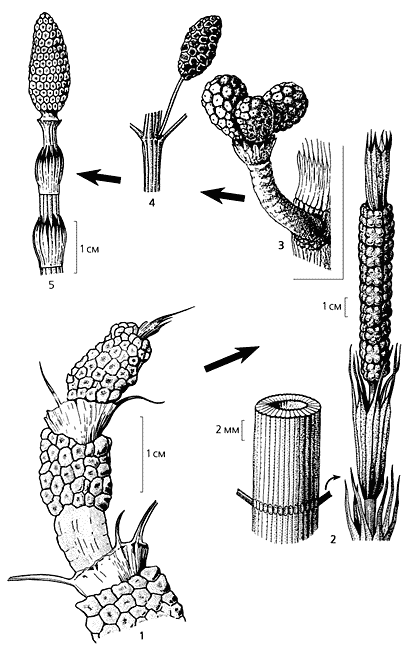
Эволюционный ряд морфогенетических изменений от пермских черновиевых до современных хвощей. Редукция генеративных органов - неизменная тенденция в этом ряду: растения древнего рода Equisetinostachys (1) имели по несколько фертильных зон; у более молодого вида Paracalamitina striata (2) осталась одна верхушечная зона, квазистробил (слева показан фрагмент стебля с веточными рубцами); затем уже появились настоящие стробилы, которые у Equisetites arenaceous (3) располагались по три на укороченных боковых побегах [7, с изменениями], а у триасового Neocalamites aff. carrerei (4) тоже на боковых ветвях, но по одному [13, с изменениями]; современный (5) хвощ полевой (Equisetum arvense) в норме несет единственный стробил на вершине осевого побега. Подобно генеративным органам, редукцию претерпели и вегетативные.Почему процветают реликты
Несмотря на то, что от громадного разнообразия палеозойских членистостебельных до настоящего времени сохранился единственный род Equisetum, современные хвощи трудно назвать исчезающими растениями. Хотя количество ныне живущих видов невелико, их заросли занимают иногда весьма обширные пространства. Нередко это бывают участки с выжженной естественной растительностью или с неплодородной глинистой землей. Тот, кому случалось неоднократно оказаться в таких местах, видимо, замечал, что со временем там появляются растения. Они могут быть довольно примитивными, но их физиологическая толерантность обеспечивает им эффективную колонизацию освободившегося пространства. Сообщества растений, которые первыми осваивают пострадавшие и опустевшие в результате разрушительных паводков или пожаров (или хозяйственной, точнее, бесхозяйственной деятельности человека), называют пионерными. Таксономическим разнообразием эти сообщества не блещут, как правило, даже очень бедны, а нередко просто состоят из единственного вида.
В средней полосе России таким видом-колонистом пионерных сообществ часто бывает хвощ полевой (Equisetum arvense), реже другие виды, например, все тот же E. hyemale. (Главные конкуренты в заселении новых пространств - безусловно покрытосеменные, особенно - травянистые формы из числа рудералов.)
В строении современного хвоща и его древних родственников много черт, связанных именно с такой экологической стратегией. В репродуктивной сфере это - специальные выросты, элатеры, которыми снабжены споры. При изменении влажности элатеры начинают сокращаться, благодаря чему сама спора движется, «переползая» с места на место. Такое передвижение весьма способствует заселению хвощами все новых территорий, при условии, что они еще не освоены другими растениями-колонистами. Элатеры были обнаружены и на спорах некоторых ископаемых членистостебельных.
Второе важное приспособление для эффективного и быстрого освоения свободных пространств - наличие горизонтальных побегов, столонов, стелющихся по поверхности субстрата или неглубоко под землей. Половой процесс, связанный с чередованием диплоидного и гаплоидного поколений (спорофита и гаметофита), у хвощей протекает довольно длительное время. И вряд ли они смогли бы процветать, если бы не вегетативное размножение. Оно с избытком компенсирует малую скорость полового размножения: из многих узлов длинных столонов развиваются новые растения, точные генетические копии родительского. Образовавшиеся заросли по сути представляют собой клон одного индивидуального хвоща. В Северной Америке таким путем размножаются некоторые стерильные гибриды хвощей и занимают обширные площади. Горизонтальные столоны также характерны для подавляющего большинства ископаемых членистостебельных, включая и каламитовые, и черновиевые.
Еще один прием, помогающий хвощам осваивать новые территории, - размножение фрагментацией, способ, который свойствен растениям водных или околоводных сообществ. При таком способе побег растения обламывается в одном из узлов, образовавшийся фрагмент переносится течением и ветром в новое место, иногда довольно далеко от материнского растения, и там укореняется.
Фрагментация известна и у некоторых ископаемых членистостебельных. Отпечатки укорененных побегов, берущих начало от фрагмента более крупного побега, обломленного в узлах, часто встречаются палеоботаникам. Но вот спороношения такого древнего хвоща практически не попадаются в ископаемом состоянии. Вероятно, причина кроется не в том, что не все органы растения оказываются в одном захоронении. Не исключено, что в тех условиях было гораздо выгоднее размножаться вегетативным путем, охватывая своими клонами значительные пространства, в том числе и непригодные для других растений.
Способность переносить неблагоприятные условия и успешно приспосабливаться к ним - качества, которые вместе с весьма выгодной и эффективной репродуктивной стратегией обеспечили хвощам долгую жизнь. Жизнь, корнями глубоко уходящую в палеозойскую эру. Теперь эти растения можно, пожалуй, назвать самыми процветающими «живыми ископаемыми».
Исследования выполнены при поддержке программы «Научные школы Российской Федерации» (проект НШ-1615. 2003.5); программы № 25 «Сравнительный анализ причинно-следственных связей и факторов глобальных биосферных перестроек в фанерозое» Президиума РАН; а также программы «Факторы эволюции биосферы» Госконтракта № 100002-251/ОН3-06/183-181/270603-908.
Литература
1. Мейен С.В. Основы палеоботаники. М., 1987.
2. Hirmer M. Handbuch der Palaeobotanik. Bd.1. Berlin. 1927.
3. Noll R. // Veroff. Museum fur Chemnitz. 2001. Bd.24. S.51-58.
4. Naugolnykh S.V. // Journ. Paleontol. 2002. V.76 (2). P.377-385.
5. Рассказова Е.С. // Тр. НИИ геологии Арктики. 1961. Вып.23-24. С.1-73.
6. Мейен С.В., Меньшикова Л.В. // Ботан. журн. 1983. Т.68. №6. C.721-729.
7. Frentzen K. // Aus Heimat. 1934. Bd.47. S.147-152.
8. Kelber K.-P., Konijnenburg-van Cittert J.H.A. van // Rev. Palaeobot. Palynol. 1998. V.100. P.1-26.
9. Вербицкая Н.Г., Радченко Г.П. Новые сибирские членистостебельные // Новые виды древних растений и беспозвоночных СССР. М., 1968. С.13-18.
10. Kashyap S.R. // J. Indian Bot. Soc. 1930. V.9. P.240-241.
11. Tschudy R.H. // Amer. J. Bot. 1939. V.26. P.744-749.
12. Zalessky M.D. // Bull. de l’Academie des Sci. de l’URSS. 1934a. №7. P. 1093-1102.
13. Владимирович В.П. // Докл. Акад. наук СССР. 1958. Т.122. №4. С.595-598.
Август 2004