№9, 2005 г.
© Алёшин В.В., Мыльников А.П., Петров Н.Б.
Дерево корненожек
В.В.Алёшин, А.П.Мыльников, Н.Б.Петров
Владимир Вениаминович Алёшин, к.б.н., в.н.с. НИИ физико-химической биологии им.А.Н.Белозерского МГУ
Александр Петрович Мыльников, д.б.н., гл.н.с. Ин-та биологии внутренних вод им.И.Д.Папанина РАН, Борок.
Николай Борисович Петров, д.б.н., зав. лаб. НИИ физико-химической биологии им.А.Н.Белозерского МГУ.Где найти амеб?
Наступивший год - юбилейный. 250 лет назад, заглянув в микроскоп, Резель фон Розенхоф открыл новую страницу в естествознании. Увиденная им живая тварь, подобно мифическому Протею, менялась на глазах, оставаясь одним существом. В цепи ее бесконечных превращений невозможно было понять план, по которому она создана творцом, не говоря о таких мелочах, как отсутствие головы и туловища, переднего и заднего конца тела, правой и левой стороны и т.п. В то время за сменой облика нельзя было увидеть того, что в амебе не меняется - структуру генов и белков, а также продуктов их работы, составляющих ее тело независимо от внешних метаморфоз.
Вскоре амеб нашли в самых разных местах - от сфагновых болот до глубин мирового океана. Они могут поселиться в крохотной лужице воды от бытового кондиционера и образовать там резервуар опасных для человека бактерий-легионелл. А могут и сами напасть на человека и вызвать амебную дизентерию, бронхит или смертельный энцефаломиелит. В последнем случае они проползут по обонятельным нервам из носовой полости в мозг купальщика подогреваемого бассейна или термального источника (амебы из обычных луж и прудов, наверное, тоже могли бы проделать такой путь, да им неуютно будет при 37°C).
Но в организме человека есть и полезные амебоидные клетки, составляющие необходимую часть нашего тела. Макрофаги и лимфоциты, похожие на маленьких амеб, пробираются между оседлыми клетками туда, где завелись микробы, где возник очаг воспаления, и там поглощают соразмерные посторонние частицы и обломки клеток или распознают чужеродные антигены. Подвижные амебоидные клетки-убийцы (n-киллеры) находят, распознают и убивают мутантные, потенциально раковые, клетки. Система внутренней безопасности многоклеточного животного подобна Протею, ее бойцы не имеют постоянной формы.
Похожа на амебу и яйцеклетка низших животных (губок, кишечнополостных), пожирающая окружающие клетки и накапливающая желток для развития будущего зародыша. А еще амебоидная подвижность незаменима в эмбриональном развитии. Многие типы клеток, которые вовсе не амебоидны в организме взрослых многоклеточных, закладываются в развитии вовсе не там, где окажутся в конце развития. Так, предшественники половых клеток у позвоночных возникают в области головы зародыша и за счет амебоидной активности переползают в зачаток половой железы. В культурах большинство типов клеток, даже происходящих из эпителиальных тканей, теряют дифференцировку и становятся похожи на амеб - выпускают ложноножки, ползают. Таким образом, все мы немного амебы. Но в амебоидности таится и смерть: вряд ли одно из самых вредоносных проявлений онкологических заболеваний - образование удаленных от первичного очага метастазов - возможно без амебоидной активности раковых клеток, внедряющихся в посторонние ткани.
Амебоидная клетка не всегда полностью бесформенная. Клетки с ограниченной свободой в изменении формы встречаются, кроме организма многоклеточных, и в природе. Например, амебы, живущие в раковинках вполне определенной формы. Одни, как фораминиферы, строят их сами, другие собирают из песчинок, как некоторые раковинные амебы наших болот, или из непереваренных остатков пищи, спикул губок и обломков домиков диатомовых водорослей, как ксенофиофореи - гигантские, в ладошку, сидячие амебоидные обитатели глубин океана. Несмотря на более или менее определенную форму домика, заключенное в него тело амебы выпускает из пор домика бессчетные ложноножки, которые образуют постоянно меняющуюся живую ловчую сеть.
Примитивны ли амебы?
У амеб нет строгого порядка в расположении разнородных деталей, который кажется нам основным признаком сложной организации, да и особых деталей в световой микроскоп не видно. Поэтому амебы с хаотически возникающими и исчезающими ложноножками переменной формы издавна создавали впечатление, что они - самые низкоорганизованные и примитивные существа на Земле, а эволюция шла “от амебы до человека”. Ранее предполагали, что наиболее примитивная из амеб - пеломикса (название происходит от греческого слова, означающего “ил”, “грязь”). Ее легко поймать, запустив руку в жидкий торф или сапропель на дне заболоченного озера. Она живет там, где почти нет кислорода: ведь черный ил и торф содержат массу органических остатков, на разложение которых (с помощью бактерий) кислород и уходит. Кислород для пеломиксы - яд. Помещенная в маленькую капельку, она, по мере растворения в ней кислорода, становится малоподвижной, собирается в комочек и погибает. В отличие от амебы протея, пеломикса не способна к дыханию, в ее клетках нет митохондрий, зато в цитоплазме живет несколько видов анаэробных бактерий. До недавних пор была в ходу гипотеза, что пеломикса - прямой потомок древнейшего эвкариота с уже оформленным ядром, но еще не вступившего в симбиоз с предками митохондрий, и что она - самое древнее “живое ископаемое”, сохраняющееся более миллиарда лет с эпохи, когда атмосфера древней Земли была восстановительная и для всех почти ее обитателей кислород был страшный яд.
Первоначально гипотезу о примитивности амеб портило только появление амебоидных клеток из жгутиковых в жизненном цикле многих видов. В самом деле, если современные амебы - самые примитивные существа на Земле, то откуда в их жизненном цикле взялись жгутиковые стадии развития? В онтогенезе многоклеточных амебоидные клетки тоже возникают из эмбриональных клеток, которые у многих видов жгутиковые. У губок жгутиковая клетка, съев много пищи, теряет жгутик, превращается в амебоцит и уходит в глубь тела, освобождая место на поверхности для голодных клеток. Внутри амебоцит может пойти целиком на пропитание яиц или передать часть питательных веществ другим клеткам, после чего возвратиться на место жгутиковой клеткой.
С точки зрения цитолога, жгутиковая клетка устроена сложнее амебы: у нее, как правило, вполне определенная форма тела: грушевидная, почковидная и т.п. Сам жгутик - сложное образование; в нем проходят упорядоченные микротрубочки: две в центре и девять пар - по периферии; всегда есть “корешковая система”, соединяющая микротрубочки жгутика с цитоскелетом клетки. Часто базальное тело жгутика и есть центр организации микротрубочек. Корешковая система отличается в разных отрядах и классах жгутиконосцев: она включает разное число микротрубочек - свободных или собранных в пучки и ленты, расходящиеся под определенными углами от базального тела жгутика. Кроме микротрубочек, в состав корешкового аппарата входят микрофиламенты. Изучив корешковый аппарат, часто удается с большой долей уверенности, даже не зная других признаков, определить систематическое положение жгутиконосца. Амебы, у которых жгутики неизвестны, очень трудно рассортировать по ультраструктурным признакам - они все кажутся на одно лицо.
Сколько существует типов эвкариотов?
В базовом учебнике по зоологии под редакцией Ю.И.Полянского (1981), по которому до сих пор учатся студенты университетов, перечислено 18 типов многоклеточных животных и пять типов одноклеточных, из которых два, “саркомастигофоры” и “книдоспоридии”, включают амебоидные формы. Нельзя сказать, что за прошедшие годы наши представления о разнообразии многоклеточных животных не изменились: описано два новых типа; многим классам, помеченным как “добавления” к “большим” типам, повысили ранг до типа; несколько “малых” типов объединили с “большими”, например, погонофор вернули в тип кольчатых червей в качестве семейства, как предлагал в 1933 г. П.В.Ушаков. Но эти реформы не идут ни в какое сравнение с перестройкой системы одноклеточных, которая сейчас полностью разрушена.
Начнем с того, что многоклеточные животные, возникнув как колония какого-то вида жгутиконосцев, родственнее этому виду (и другим его одноклеточным потомкам), чем остальным одноклеточным, дожившим до наших дней, и таким образом представляют собой только одну из веточек разветвленного филогенетического дерева одноклеточных. Этот чисто логический вывод идет вразрез с установившейся практикой деления живых существ на одноклеточных и многоклеточных.
С помощью электронного микроскопа выяснили, что ультраструктура одноклеточных (главным образом жгутиковых форм) неодинакова. В результате было предложено от полудюжины до полутора десятков царств “одноклеточных”. Имея перед глазами пример перекройки системы жгутиконосцев, все протистологи соглашались, что амебы - временное собрание неродственных организмов, с которым до поры приходится мириться ввиду отсутствия надежного метода анализа. И только в последние годы многие амебоидные организмы нашли свое место на филогенетическом дереве.
Первые этапы ревизии амеб
Современная ревизия амеб началась с уничтожения “книдоспоридий” (они же миксоспоридии) как типа “одноклеточных”. Сначала показали, что два номинальных класса этого типа - только разные стадии в жизненном цикле одного и того же вида, паразитирующие, соответственно, в позвоночных и кольчатых червях. А затем анализ генов рибосомной РНК и генов семейства Hox, которые встречаются только у многоклеточных животных (управляют эмбриональным развитием) и у миксоспоридий (функция неизвестна), доказал, что последние - это упростившиеся многоклеточные, утратившие все без исключения морфологические и эмбриональные признаки Metazoa [1].
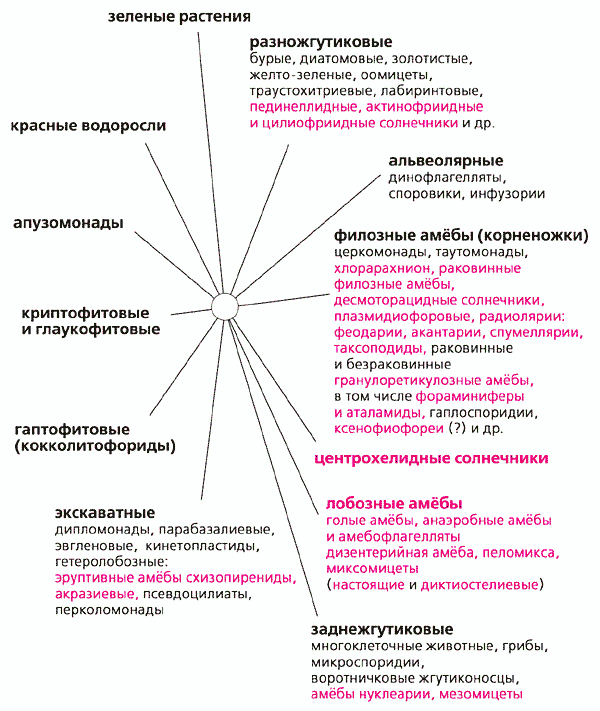
На втором этапе, который не завершен и продлится еще долго, началось распознавание отдельных амебоидных представителей в различных группах жгутиконосцев, недавно утративших жгутики, или даже и не утративших, а просто большую часть жизненного цикла проводящих как амебы. Систематическое положение некоторых из них не вызывало сомнения и у протистологов прошлого. Таков непатогенный кишечный комменсал Dientamoeba - амебоидный представитель парабазалий [2], остальные виды которых - жгутиконосцы, как всем известная трихомонада или гипермастигины из кишечника термитов, участвующие в усвоении клетчатки. Другой пример - пединеллидные жгутиконосцы; среди них есть фотосинтезирующие и гетеротрофные виды, выпускающие ловчую сеть из ложноножек. Их родство никогда не вызывало сомнений ввиду очевидного морфологического сходства. Амебоидные (солнечникообразные) пединеллиды, которых называют цилиофриидами, как правило, сохраняют жгутик, который соответствует переднему жгутику разножгутиковых (Heterokonta) [3].
Корненожки - амебы с тонкими ложноножками
Намного сложнее вопрос о положении видов, включенных в учебниках в класс “саркодовые” как “корненожки”, “радиолярии” и “солнечники”. Первый существенный прорыв в определении родственных связей этих организмов произошел в 1995 г. [4]. Тогда при сравнении генов 18S рРНК (рибосомальной РНК) обнаружилась близость обычных почвенных гетеротрофных раковинных филозных амеб * (жгутиковые стадии в жизненном цикле не обнаружены) с необычной зеленой амебой хлорарахнионом (в переводе с греческого - зеленая паутина). Жизненный цикл хлорарахниона начинается с двужгутиковой клетки - зооспоры, которая оседает на дно и начинает расти. По мере роста она выпускает длинные тонкие выросты, делится, но клетки не расходятся полностью, а остаются связаны тонкими нитями в сложную сеть. От нитей грибницы она отличается тем, что в ней отчетливо видны клетки и связывающие их перемычки и нитевидные ложноножки. Хлорарахнион прославился тем, что его клетка устроена как матрешка. Его хлоропласты, казавшиеся не примечательными до тех пор, пока их не изучили с помощью электронного микроскопа, в действительности оказались эвкариотическими симбионтами, родственными зеленым водорослям. В них сохранился рудимент ядра - нуклеоморф с генами, больше похожими на гены зеленых водорослей, чем на гомологичные гены ядра хлорарахниона. Например, рибосом в клетке хлорарахниона четыре типа: первый - цитоплазматические, их компоненты кодирует ядро; второй - митохондриальные (прокариотического типа), их компоненты кодируют гены митохондриального генома; третий - хлоропластные эвкариотического типа, их кодирует нуклеоморф, и похожи они на цитоплазматические рибосомы зеленых водорослей; четвертый - хлоропластные прокариотического типа, кодируются хлоропластной ДНК и похожи на рибосомы цианобактерий, так же как хлоропластные рибосомы зеленых растений.
* У этих амеб тонкие ложноножки, в которых нет микротрубочек, а есть только микрофиламенты.Следующее открытие произошло через пару лет [5]. Оказалось, что гены рРНК филозных амеб и хлорарахниона похожи на гены рРНК гетеротрофных жгутиконосцев-церкомонад. У некоторых жгутиконосцев рРНК похожа на амебную больше, чем на рРНК других видов жгутиконосцев. Церкомонады - это очень многочисленные, разнообразные и вездесущие организмы [6-8]. Если подцепить ногтем немного почвы - в ней наверняка найдется несколько видов церкомонад. Раньше на них не обращали особого внимания, поскольку среди них не значилось вредных или полезных видов, да и вообще виды церкомонад с трудом отличали. Церкомонадам свойственна высокая морфологическая изменчивость: то это грушевидная двужгутиковая клетка (с неравными жгутиками), то на заднем полюсе она начинает выпускать ложноножки - тонкие филоподии или широкие лобоподии (в зависимости от вида) и превращается в амебофлагелляту, может осесть на дно, распластаться и образовать сеть клеток (плазмодий), наподобие хлорарахниона, только небольшую; потом клетки могут отсоединиться и вновь стать грушевидными жгутиконосцами без всяких ложноножек. Видимо, большую часть жизни церкомонады проводят как амебы с малоподвижными жгутиками. Однако это стало понятно совсем недавно. Увидеть что-то в микроскоп можно только при хорошем освещении, но церкомонады избегают света. Как только природный образец с церкомонадами в виде амеб помещают на предметное стекло, за несколько секунд они активизируют жгутики и отсоединяются от субстрата, пытаясь вновь уйти в темноту, и в поле зрения оказываются только беспорядочно плавающие жгутиконосцы.Следующая группа родственников церкомонад и филозных амеб - плазмодиофоры. Эти хозяйственно важные внутриклеточные паразиты растений вызывают килу капусты, порошистую паршу картофеля и другие заболевания. Плазмодий, т.е. многоядерная клетка плазмодиофоровых, лежит в паразитофорной вакуоли и на первых порах стимулирует гипертрофированный рост и ускоренное деление зараженных клеток, вызывая опухолевидное разрастание пораженного участка. На втором этапе цитоплазма хозяйских клеток постепенно исчезает, и оболочка заполняется массой одноядерных спор (цист) паразита. Жизненный цикл плазмодиофоровых, как и многих церкомонад, весьма сложный. У возбудителя килы капусты (и других крестоцветных) Plasmodiophora brassica он включает: одноядерные амебы; двуядерные амебы; два поколения многоядерных амеб-плазмодиев, из которых гаплоидное паразитирует в корневых волосках, а гетерокариотическое - в клетках гипокотиля; расщепление плазмодиев на спорангии; половой процесс; два типа спор (цист); двужгутиковые зооспоры. В течение жизненного цикла паразит дважды покидает хозяина, чтобы пройти часть развития в почве. Долгое время плазмодиофор относили за их плазмодий к миксомицетам, когда установили, что они не грибы.
Другая группа паразитических организмов - гаплоспоридии, имеет намного более длинный послужной список перемещения по системе простейших. С кем их только не сближали: с микроспоридиями, споровиками, миксомицетами, дизентерийными амебами. Однако родство с церкомонадами, раковинными филозными амебами и плазмодиофорами, установленное по генам 18S рРНК, стало полной неожиданностью. Гаплоспоридии - облигатные тканевые паразиты беспозвоночных, полностью лишенные жгутиковых стадий. Их вегетативное тело представляет одноядерную амебу или дикариотический плазмодий. Созревший плазмодий разделяется на многочисленные одноядерные споробласты, которые, как считается, сливаются попарно и каждый образует спору совершенно необычного строения, состоящую из двух частей. Внешняя похожа на шкатулку с чашевидным основанием и крышечкой. Она образована толстым слоем эписпорной цитоплазмы с различными органеллами (митохондриями, вакуолями, эндоплазматическим ретикулюмом), но без ядра. Внутреннее пространство шкатулки занимает одноядерная клетка, окруженная плазмалеммой.
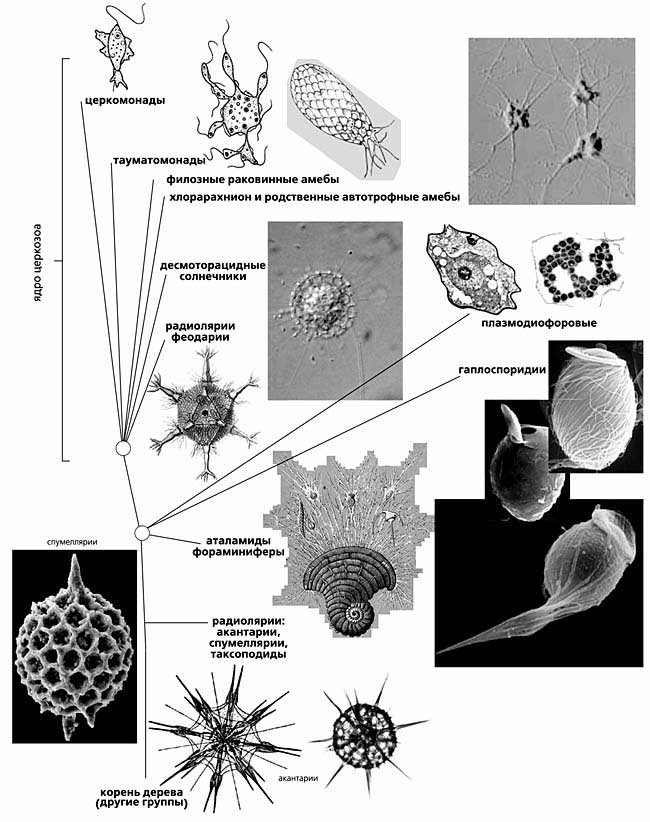
Супертаксон церкомонады
Итак, амебофлагелляты-церкомонады, вместе с их близкими и далекими родственниками, составляют очень крупную группу одноклеточных. Истинный ее размер стал понятен только сейчас [9]. Этот гигантский монофилетический таксон-агломерат вбирает в себя большинство амебоидных организмов с филоподиями и ретикулоподиями (тонкими анастомозирующими ложноножками, образующими ловчую сеть вокруг клетки, например, у фораминифер). Одни представители этого таксона - обычные жгутиконосцы, другие - амебы, никогда не имеющие жгутиков, а многие виды совмещают в сложном жизненном цикле жгутиковые и амебоидные стадии. В таксоне много лучистых одноклеточных - радиолярий и солнечников, образующих несколько эволюционно независимых ветвей. Например, десмоторацидные солнечники - изящные округлые клетки, сидящие на длинной ножке. Они заключены в органическую капсулу, из окошек которой выпускают во все стороны тонкие аксоподии. На аксоподиях заметны светопреломляющие гранулы (экструсомы) - признак любого солнечника. Это стрекательные органеллы. При соприкосновении с добычей (мелким жгутиконосцем) составляющие их белки мгновенно меняют конформацию, гранула перестраивается в тончайшую длинную белковую нить, которая убивает и запутывает жертву. Деление солнечника неравномерное: одна дочерняя клетка - амебоидная, она остается в домике, а другая - двужгутиковая - уплывает заселять новые места. В дальнейшем она отбросит жгутики и превратится в ползающую в илу амебу, которая в подходящем месте прикрепляется и образует ажурный домик на ножке.
Установление родства радиолярий и раковинных амеб позволяет представить, как возникла их организация. Известно, что клетка радиолярии подразделяется на два отсека: центральная часть ее заключена в ажурную капсулу из сросшихся минеральных игл и пластинок - развитого внутриклеточного скелета; вне капсулы находится обычно большая по объему и сильно вакуолизированная часть цитоплазмы. Легко представить себе, что она - производное ложноножек, наподобие тех, что выпускают фораминиферы и десмоторацидные солнечники из окошек домика, но которые слились в обширную сеть, обволакивающую раковину толстым рыхлым слоем живого вещества. Из такой гипотетической схемы морфологических превращений следует, что планктонные организмы - радиолярии - возникли из бентосных раковинных амеб. Причем не менее чем дважды. Одна, более ранняя ветвь, включает акантарий, спумеллярий и наиболее специализированных, потерявших внутриклеточную капсулу билатерально-симметричных таксоподиальных радиолярий. Другая - феодарии, имеющие полый скелет из кремнезема, - возникла независимо в ходе радиации линий, составляющих ядро таксона корненожек. Кроме внекапсулярной цитоплазмы (слившихся ложноножек), у радиолярий есть и аксоподии. Радиолярии используют их, в отличие от солнечников, только для парения и передвижения; на них не бывает стрекательных органелл для поимки крупной добычи.
В каких других эволюционных линиях возникло много амебоидных организмов? Рассмотрим филогенетическое дерево эвкариотов в целом. Если исключить из него некоторые редкие и малоизвестные виды, то все эвкариоты распределятся по 12 монофилетическим супертаксонам. Среди них есть относительно малочисленные, включающие какую-то сотню видов, другие - гигантские по числу видов. Например, супертаксон заднежгутиковых объединяет многоклеточных животных, царство грибов, воротничковых жгутиконосцев и некоторые другие организмы. Современный уровень знаний не позволяет пока установить, в каких родственных отношениях между собой находятся эти 12 супертаксонов. В половине их имеются более или менее многочисленные группы амеб. Один из них мы только что рассмотрели, о других поговорим в следующий раз.
Литература
1. Алёшин В.В., Петров Н.Б. Регресс в эволюции многоклеточных животных // Природа. 2001. №7. С.62-70.
2. Silberman J.D. et al. // Mol. Biochem. Parasitol. 1996. V.76. №1-2. P.311-314.
3. Микрюков К.А. Центрохелидные солнечники. М., 2002.
4. Bhattacharya D. et al. // J. Eukaryot. Microbiol. 1995. V.42. №1. P.65-69.
5. Cavalier-Smith T., Chao E.E. // Arch. Protistenkd. 1996/97. V.147. P.227-236.
6. Мыльников А.П. Класс Cercomonadea // Протисты. Ч.1. СПб., 2000. С.411-417.
7. Cavalier-Smith T., Chao E.E. // Protist. 2003. V.154. №3-4. P.341-358.
8. Myl’nikov A.P., Karpov S.A. // Protistology. 2004. V.3. №4. P.201-217.
9. Nikolaev S.I. et al. // Proc. Natl. Acad. Sci. USA. 2004. V.101. №21. P.8066-8071.