№ 10, 2000 г.
№ 10, 2000 г.
№ 10, 2000 г.
№ 10, 2000 г.
© А.С. Антонов
Растения и животные - “живые ископаемые”
А.С. Антонов
Андрей Сергеевич Антонов, доктор биологических наук, профессор,
заведующий отделом эволюционной биохимии
научно-исследовательского института физико-химической биологии им.А.Н.Белозерского
Московского государственного университета.В 1939 г. ихтиолог Дж.Смит описал странную рыбу, которую по всем признакам следовало бы отнести к группе кистеперых, давным-давно вымерших, как полагали палеонтологи. Сегодня таких рыб обычно называют латимериями, или целакантами. Полностью исследовать пойманный экземпляр тогда не смогли. В руки зоологов попала одна шкура.
Прошло 13 лет, прежде чем у Коморских о-вов поймали вторую латимерию. Рыбу удалось сохранить целиком - это оказался представитель группы, которая, как считалось, окончательно вымерла около 70 млн лет назад. Все выводы, сделанные Смитом на основании изучения первого неполного экземпляра, подтвердились. Журналисты тут же окрестили рыбу “живым ископаемым”. На некоторое время ихтиологи и палеонтологи заняли первые полосы газет и журналов - рассуждать об истории развития жизни на Земле стало модным.
Латимерия - отнюдь не единственное “живое ископаемое” в мире животных. Интерес к ней был вызван тем, что она оказалась первой подобной “ископаемой” среди казалось бы прекрасно изученных позвоночных животных.
Не менее известный представитель “живых ископаемых” - моллюск неопилина. Первоначально, в 1940 г., класс моллюсков, включающий неопилин, был выделен в результате изучения ископаемых остатков. Однако через 12 лет живых неопилин поймали в Тихом океане! В газетах и научно-популярных журналах снова было много шума, отголоски которого слышны до сих пор: все спорят, живет ли в озере Лох-Несс плезиозавр, в Конго - птеродактили, и т.д. К сожалению, в руки ученых “живые ископаемые” попадают редко.
Вообще-то как среди животных, так и среди растений древние виды довольно широко распространены. Обыкновенный щитень, обитающий в чистых лужах экологически благополучных областей нашей страны, по внешним признакам как две капли воды похож на своих далеких предков, живших около 200 млн лет назад.
И “живые ископаемые”, и древние виды являют собой поразительный пример того, как внешние признаки некоторых организмов, их фенотипы, могут сохраняться неизменными многие десятки и сотни миллионов лет. Означает ли это, что их генетические программы (генотипы) также остаются законсервированными на столь же долгий срок? Если так, то как им удается “гасить” возникающие мутации и не давать им проявляться в фенотипах?
Ответ на этот вопрос получили лишь после того, как молекулярные биологи научились определять структуру так называемых семантид (ДНК, РНК и белков), участвующих в хранении и передаче генетической информации. Как известно, все три класса семантид - полимеры, построенные из простых мономеров. По ряду косвенных признаков уже в первой половине ХХ в. ученые предполагали высокую специфичность строения белков: им даже отводили ведущую роль в передаче наследственных признаков организмов. Значительно позже на сцену вышли нуклеиновые кислоты - ДНК и РНК.
Уже первые опыты, поставленные в 50 - начале 60-х годов Э.Чаргаффом, А.Н.Белозерским и их сотрудниками, показали, что количество типов мономеров и их взаиморасположение в ДНК и РНК у разных видов индивидуально, видоспецифично. Позднее выяснили, что по этому признаку и “живые ископаемые”, и просто древние виды ничем не отличаются от более молодых. Семантиды у них эволюционировали по тем же законам, что и у самых обычных видов, фенотипы которых приобретали все новые черты в ходе эволюции, но мутации в геномах “живых ископаемых” и древних видов не изменяли их внешних черт. Иными словами, они находились под влиянием мощного стабилизируюшего отбора, действовавшего на уровне фенотипов.
Факт, что ни говори, удивительный: внешние признаки организмов почти не менялись, а все их “внутреннее содержание” (семантиды и эписемантиды - “вторичные вещества”) продолжало эволюционировать. Можно утверждать, что биохимия современной латимерии отличается от биохимии вымершей кистеперой рыбы не меньше, чем, допустим, вымершей латимерии и современного окуня. То же можно сказать и о неопилине, и о щитне, и о сотнях других видов. Конечно, сегодня нельзя изучить структуру семантид у давно вымерших животных и растений путем прямого анализа - ведь эти соединения в ископаемых остатках давным-давно разрушились. Однако современная молекулярная филогенетика располагает целым набором методов реконструкции строения отдельных генов и других участков ДНК у давно вымерших животных и растений путем сопоставления их строения у ныне живущих форм.
В мире растений также известно достаточно много древних видов. Среди высших, семенных растений к ним относится, например, гинкго. В далеком прошлом в гинкговых лесах паслись динозавры. Если же мы сравним фенотипы ископаемых и современных растений гинкго, то вряд ли обнаружим в них сколько-нибудь существенные различия.
Совсем другой результат получится, если сопоставить строение семантид у гинкго и его ближайших родственников, а затем использовать эти данные для реконструкции событий молекулярной эволюции семантид у растений. Из построенного таким образом молекулярного филогенетического древа со всей очевидностью следует, что ныне живущие гинкго - представители давно обособившейся линии эволюционного развития голосеменных растений, и в их ДНК, РНК и белках за сотни миллионов лет произошли большие изменения.
Таким образом, можно утверждать, что эволюция генотипов и фенотипов происходила отнюдь не параллельно. Иногда изменения фенотипов заметно отставали от таковых в генотипах, в других случаях они менялись более или менее согласованно, а в третьих фенотипы менялись существенно быстрее, чем генотипы. Так, в оз.Виктория в Африке существует множество хорошо отличимых видов рыб-цихлид, генотипы которых почти одинаковы. Видимо, все они произошли от какого-то одного предка, попавшего в этот водоем вскоре после его образования и послужившего основой для бурного видообразования. Сходные данные получены при изучении некоторых групп рыб из оз.Байкал. В этих случаях “генетическая цена” образования новых видов оказалась небольшой.
Аналогичные примеры известны и в мире растений: попав на какой-нибудь остров или архипелаг, некий вид становится родоначальником целого куста (рода или даже небольшого семейства) хорошо различимых видов, но этот процесс обычно не сопровождается накоплением пропорциональных изменений в их генотипах.
Приведенные примеры известны биологам давно. Виды растений и животных с неизмененным в ходе эволюции фенотипом находили в основном в эволюционно древних группах. Вероятность существования таких форм среди покрытосеменных растений, до недавнего времени (по данным палеонтологии) считавшихся относительно эволюционно молодыми группами, была небольшой.
Мир покрытосеменных изучен ботаниками неплохо (впрочем, не лучше, чем рыбы к моменту описания латимерии). Само их возникновение относили к началу мелового периода (140-125 млн лет назад). Сопоставив фенотипы современных и ископаемых покрытосеменных, большинство ботаников пришло к выводу, что самые древние среди них - магнолиевые.
Впервые в молодости покрытосеменных усомнился английский биохимик Д. Боултер с коллегами. Схватку Боултера и известного ботаника А. Кронквиста мне довелось наблюдать на одном из симпозиумов Всемирного ботанического конгресса в Ленинграде. Боултер занимался тогда сравнительным изучением строения фермента цитохрома с.
В то время уже получила широкое хождение так называемая гипотеза молекулярных часов, предложенная Э. Цукеркандлем и Л. Полингом. Согласно этой гипотезе, для оценки времени расхождения двух линий эволюционного развития достаточно установить количественные различия в строении их семантид. Такой подход и использовал Боултер относительно покрытосеменных и получил удивительный результат: по его данным, покрытосеменные отделились от других, даже самых близких линий (например, голосеменных) не 140-125, а около 450 млн лет назад.
В дискуссии по докладу Кронквист один за одним наносил убийственные удары по построениям Боултера.
Уверен ли автор в том, что аминокислотные последовательности белков определены правильно? Ведь число наблюдаемых изменений невелико, и всякая ошибка скажется на топологии получаемых деревьев.
Как можно использовать для расчета времени разделения данные о скорости эволюции цитохромов, полученные для совсем другой группы, - грибов? Биологам известно, что скорость изменений может быть неодинаковой в разных линиях эволюционного развития.
Уверен ли автор в правильности выбора метода реконструкции филогенетического древа? Ведь использованный им метод основан на гипотезе молекулярных часов, которая сама по себе требует доказательств.
Неужели он всерьез считает, что устоявшиеся взгляды можно столь радикально менять, проанализировав всего десяток-другой белков? В самом деле, ботаники для своих филогенетических построений сопоставляют тысячи видов, а здесь объем выборки явно маловат.
В какой мере эволюция одного белка отражает эволюцию организмов? Действительно, обычно один белок - продукт всего одного гена (впрочем, есть и сложные белки, построенные из нескольких субъединиц, каждая из которых кодируется своим, отличающимся по структуре геном), а в генотипах растений генов десятки тысяч.
Четких ответов на эти вопросы Боултер тогда не дал, но доклад запомнился: было ясно, что он предполагает совсем новый путь изучения эволюции растений.
В это же время в лаборатории Белозерского в МГУ искали другой способ решения тех же проблем: с конца 50-х годов в ней применялись все новые методы сравнительного изучения строения ДНК, которые поставляла бурно развивающаяся молекулярная биология. Использование их позволяло шаг за шагом продвигаться к пониманию молекулярной эволюции генотипов растений. Об этих работах мне уже приходилось писать в журнале “Природа” [1].
Наконец, где-то в конце 70 - начале 80-х годов молекулярные биологи разработали удобные методы определения нуклеотидной последовательности в ДНК и РНК. Вместо трудоемкой работы по расшифровке строения белков гораздо проще оказалось анализировать структуру кодирующих их генов. Изучение молекулярной эволюции генотипов растений пошло быстрее. Появилась возможность либо опровергнуть, либо подтвердить достоверность предварительных выводов Боултера.
Конечно, тогда речь не шла об определении строения всей ДНК растительной клетки - решать эту задачу начали лишь в конце 90-х годов. В последних номерах “Nature” уже вышли статьи с описанием полной структуры ДНК двух хромосом покрытосеменного растения - арабидопсиса. До полной расшифровки строения его генома остались считанные годы, а на очереди - геномы риса, кукурузы, пшеницы и других растений.
Первоначальная задача была скромнее: на уровне ДНК (или же ее комплементарных копий, молекул РНК) исследовать эволюцию отдельных генов семенных растений и попытаться сопоставить полученные данные с представлениями ботаников об эволюции этой группы. Первыми к решению этой задачи приступили японские исследователи [2]. К сожалению, при построении филогенетических древес изученной ими так называемой 5S рибосомной РНК они исходили из гипотезы молекулярных часов и к тому же применили простой (так называемый кластерный) метод, что делало их выводы недостаточно надежными.
Как известно, на ошибках учатся, и это удобно, если ошибки делает кто-то другой. Мы использовали иной подход: прежде всего последовательно выбирали такие методы реконструкции филогенетических древес, топология которых в минимальной степени зависела от недостаточно обоснованных гипотез о механизмах молекулярной эволюции. Существенную помощь на этом этапе нам оказали новосибирские ученые во главе с В.А.Ратнером, которые давно занимались моделированием процессов молекулярной эволюции. Было решено проследить эволюционный путь сначала одного, а потом сразу нескольких генов рибосомных РНК [3]. Не теряли времени и наши зарубежные коллеги: в 1989 г., помимо нашей лаборатории [4] ряд зарубежных опубликовал первые результаты изучения эволюции некоторых генов семенных растений [5].
Таким образом, к началу 90-х годов мы уже имели представление о самых общих закономерностях молекулярной эволюции покрытосеменных. Как мне представляется, наиболее важным был сделанный нами вывод о монофилии (т.е. о происхождении всей группы от какого-то одного гипотетического предка) как голо-, так и покрытосеменных растений. К тому же вскрылось еще одно важное обстоятельство: разделение этих линий произошло до того, как сформировались все ныне существующие (и вымершие) группы голосеменных растений. По уточненным данным, событие произошло 360 млн лет назад [6].
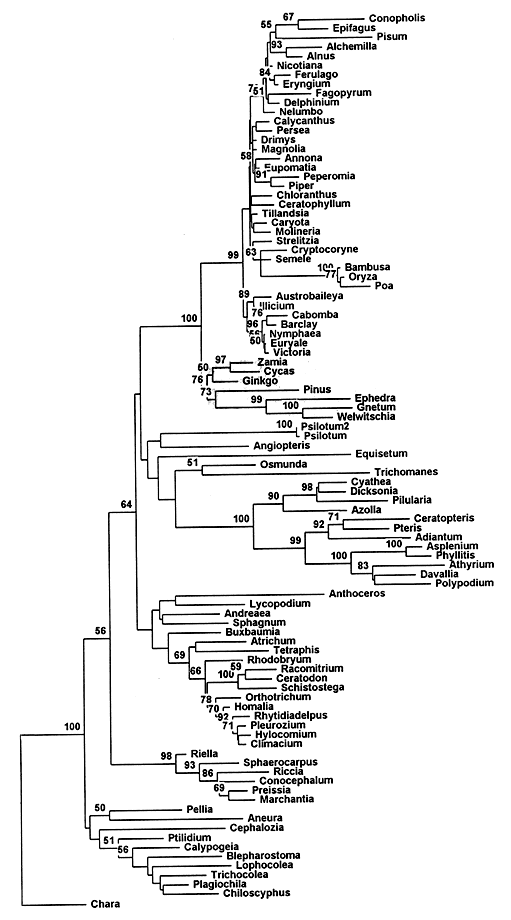
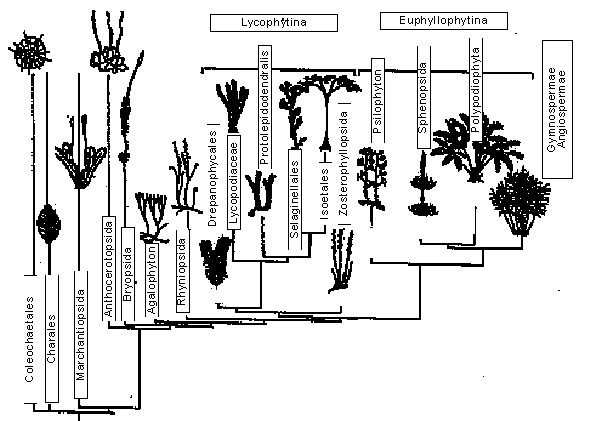
Из палеоботаники известно, что все основные группы голосеменных разошлись в своем эволюционном развитии очень давно и большинство из них существует на Земле сотни миллионов лет (вспомните гинкго!). Значит вывод мог быть только один: “возраст” покрытосеменных гораздо больше, чем это следовало из данных палеонтологии. Тем самым подтверждалась еретическая гипотеза Боултера, а ботаникам пришлось задуматься, стоит ли так безоглядно доверять своим коллегам-палеоботаникам при определении времени расхождения эволюционных ветвей растений.Филогенетические древа. Сверху - один из вариантов, построенных по молекулярным признакам генотипов растений. Числа показывают вероятность (в %) происхождения данной группы видов от единого предка.
Снизу - схема, отражающая представления ботаников об истории становления основных наземных растений и их взаимосвязи с зелеными водорослями. Указаны как вымершие, так и ныне существующие группы. Видно, что хотя ботаники и молекулярные биологи по-разному оценивают положение некоторых групп, например антоцеротовых, в целом два дерева очень схожи.Впрочем, претензий к палеонтологам быть не может: не находили и не находят они среди самых древних ископаемых растений ничего похожего на современных покрытосеменных. Разных голосеменных - сколько угодно, а покрытосеменные никаких материальных следов своего существования на Земле с момента отделения от общего предка с голосеменными не оставили. Правда, на последнем Московском совещании по филогении растений (январь 2000 г.) профессор МГУ Н.Р.Меликян, сравнив строение пыльцы современных покрытосеменных и ископаемых спор так называемых семенных папоротников, высказала гипотезу об их частичном сходстве. А это как раз та группа, которую считают наиболее вероятным предком покрытосеменных. Этот результат обнадеживает: со временем палеоботаники, по-новому взглянув на ископаемые растения, которых они дружно считали предками современных голосеменных, либо обнаружив новые места, где сохранились их ископаемые остатки, смогут все же выделить группу, с которой в меловом периоде началась бурная эволюция современных покрытосеменных растений.
При всей важности полученных результатов, остается нерешенным один существенный вопрос: почему среди голосеменных виды с замедленной эволюцией фенотипов - явление частое, а среди покрытосеменных они встречаются существенно реже?
Интересно, что данные, полученные геносистематиками, и представления ботаников о ранних стадиях эволюции покрытосеменных в некоторых деталях расходятся. Анализ эволюции самых разных генов указывает, что среди ныне существующих покрытосеменных первыми от общего ствола отделились виды из семейств амборелиевых, нимфейных, иллициевых и ряда других, известных под сборным названием “палеотравы”. Справедливости ради отметим, что в их строении ботаники давно усматривали много примитивных черт, но часть из них все же относили к более продвинутым таксонам. Между тем, по результатам молекулярного анализа, лишь после формирования палеотрав от ствола покрытосеменных отделились линии, ведущие к современным магнолиидам и однодольным растениям, а в конечном счете - и к высшим двудольным. Вот эти-то группы и представлены в ископаемом материале, и именно среди них и встречаются современники кистеперых рыб. Назвать их “живыми ископаемыми” мы все-таки не можем: палеонтологи и ботаники описали их очень давно и дружно отнесли к древнейшим покрытосеменным.
Остается надеяться, что в древних слоях Земли со временем будут найдены и ископаемые палеотравы, и это подтвердит выводы геносистематиков о порядке расхождения основных ветвей развития покрытосеменных.
А.Л.Тахтаджян отмечал, что скорее всего “первичный центр происхождения магнолиофитов находился далеко от известных нам мест их фоссилизации (т.е. мест нахождения их ископаемых остатков. - А.А.)” [7]. Он же высказал предположение, что таким центром могли послужить некие острова, располагавшиеся между Гондваной и Лавразией.
В самом деле, обидно, что украшение наших рек и озер, желтые кубышки и белые водяные лилии - Иваны, не помнящие родства, а фасонистые магнолии могут указать своих родичей вплоть до мелового периода.
С другой стороны, можно пофантазировать, что где-нибудь (скорее всего в весьма ограниченном районе тропиков, прямо связанном своей геологической историей с загадочными островами) ботаники обнаружат покрытосеменные растения, которые можно будет соотнести с ископаемыми остатками домелового возраста. Только тогда с полным основанием мы скажем, что и среди покрытосеменных растений есть “живые ископаемые”.
Некоторые находки ботаников допускают такую возможность: нашли же они на о-вах Фиджи удивительное растение - дегенерию, на Мадагаскаре - тахтаджанию, а на Новой Каледонии - амборелу, которую многие молекулярные биологи ставят сегодня у самого корня покрытосеменных растений. Только вот ископаемых, домеловых остатков, которые соотносились бы с этими морфологически примитивными растениями, обнаружить пока не удалось.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Проект 96-15-97970.
Литература
1 Антонов А.С. Происхождение основных групп наземных растений // Природа. 1997. №10. С.55-63; Он же. Геномика и геносистематика // Там же. 1999. №6. С.19-26.
2 Hori H., Lim B.-L., Osawa S. // Proc. Natl. Acad. Sci. USA. 1985. V.82. P.820-823.
3 Troitsky A.V. et al. // J. Mol. Evol. 1991. V.32. P.253-261.
4 Рахимова Г. М. и др. // Молекуляр. биология. 1989. Т.23. С.83-841.
5 Martin W. et al. // Nature. 1989. V.339. P.46-48; Wolfe K.H. et al. // Proc. Natl. Acad. Sci. USA. 1989. V.86. P.6201-6205.
6 Goremykin V. et al. // Plant Syst. Evol. 1997. V.206. P.337-351.
 Октябрь 2000 |