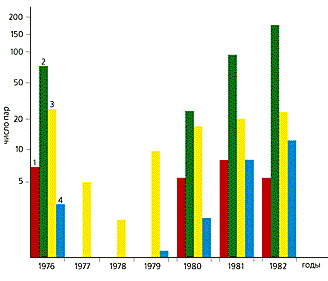№10, 2002 г.
№10, 2002 г.
№10, 2002 г.
№10, 2002 г.
© Ф.Б.Чернявский
Лемминговые циклы
Ф.Б. Чернявский
Феликс Борисович Чернявский, доктор биологических наук,
директор Института биологических проблем Севера ДВО РАН (Магадан),
заведующий лабораторией экологии млекопитающих.Взгляды зоологов на природу периодических колебаний численности леммингов в тундровой зоне Голарктики были изложены на страницах журнала “Природа” более 30 лет назад [1]. Но тогда механизм этого довольно хорошо изученного явления оставался загадкой. Все прошедшие годы небольшие зверьки из подсемейства полевок, играющие важную роль в жизни тундровых биоценозов, продолжали интересовать специалистов многих стран [2]. Лемминговые циклы — один из частных случаев общей экологической проблемы, которая уже несколько десятилетий служит предметом острых дискуссий.
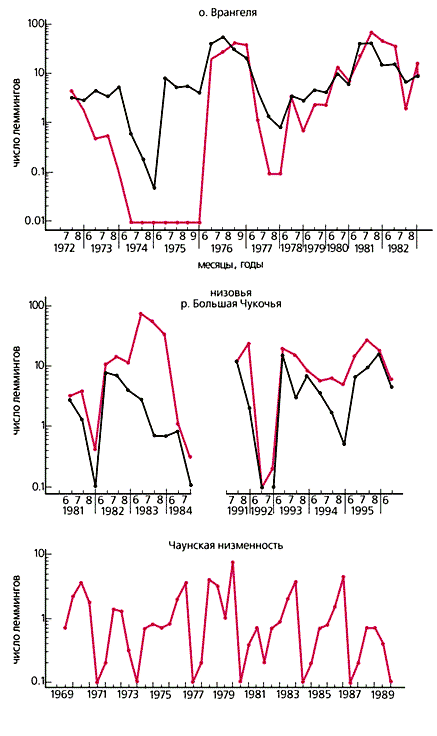
Лемминг копытный. Фото автора В 1972 г. мы начали долговременную программу по изучению динамики численности леммингов сибирского (Lemmus sibiricus) и копытного (Dicrostonyx torquatus) на о.Врангеля, в Чаунской и Колымской низменностях. Сегодня хорошо известно, что правильная периодичность пиков их численности — четкая закономерность, свойственная всей тундровой зоне от Скандинавии до Северной Канады. Пики, как правило, отделены друг от друга промежутками в три-четыре года и происходят синхронно у обоих видов грызунов.
Сравнение лемминговых циклов в разных подзонах тундры в пределах северо-востока Сибири показало, что их структура подвержена географической изменчивости. Наиболее четкая картина наблюдается в сравнительно малопродуктивной арктической экосистеме, но имеющей достаточно полный набор хищников и условий для обитания обоих видов тундровых леммингов. Так, на о.Врангеля амплитуда годовых колебаний численности и длина периодов между ними, судя по нашим данным, возрастает с увеличением широты. Участие копытного лемминга в цикле зависит от наличия подходящей для него ниши в экосистеме. В южной, сравнительно высокопродуктивной тундре с неполным набором хищников циклы имеют малую амплитуду. Это говорит о том, что структура тундровой экосистемы играет существенную роль в характере леммингового цикла.Демография
Основные демографические параметры, от которых зависит численность любой популяции — это показатели размножения, смертности и половозрастной структуры. Продолжительные наблюдения за лемминговыми популяциями на о.Врангеля (1972—1982) и в Колымской низменности (1981—1996) позволили получить довольно четкую демографическую картину.
Давно известно, что лемминги способны размножаться под снегом. По возрастному составу выборок из популяций обоих видов грызунов, отловленных в июне, видно, что весной эта уникальная особенность проявляется на всех стадиях цикла, но с разной интенсивностью: самая высокая в годы роста численности (у взрослых самок не менее пяти-шести подснежных пометов), самая низкая — в годы спада. Оба вида леммингов регулярно приносят выводки в январе—апреле, хотя на всех фазах цикла активнее размножаются в марте—апреле. Репродукция разной степени интенсивности происходит в марте — апреле. Это подтверждает точку зрения, что подснежное размножение тундровых зверьков, несмотря на пониженную плодовитость самок, приводит к росту популяций [3].
Летом в разных районах тундры взрослые самки приносят два-три выводка. Иногда один-два помета дают и молодые самки-сеголетки. Продолжительность летней репродукции, точнее, сроки ее окончания, сильно колеблется в зависимости от плотности популяции. Особенно рано заканчивается размножение, когда численность грызунов максимальна. Так, на о.Врангеля в фазе пика (1976) у сибирских леммингов оно прекратилось в конце июля, а у копытных еще раньше — в середине этого месяца. Повышенная плотность популяции приводит также к нарушению репродуктивных функций: резко снижается вес семенников, прекращается сперматогенез у самцов и смещается эстральный цикл у самок. В зависимости от фазы цикла меняется и интенсивность летнего размножения. На о.Врангеля она сильнее (наивысшая доля самок и самцов, участвующих в размножении) в фазе роста численности. На фазах спада и депрессии, несмотря на значительную продолжительность летнего размножения, репродуктивная активность сравнительно низкая [3].
Таким образом, летняя плодовитость у сибирского и копытного леммингов снижена на фазах спада и депрессии, заметно увеличена в фазе роста и предельно высока при максимуме численности. Динамика роста лемминговых популяций свидетельствует о высокой смертности зверьков зимой, после фазы пика, а также повышенной ювенильной смертности (гибели уже родившегося молодняка в гнездах) летом, в фазе пика. В переуплотненных популяциях грызунов замедлено половое созревание молодняка, в фазе роста изменяется возрастная структура популяции (омоложение), а у копытных леммингов соотношение полов сдвигается в пользу самок.
Итак, при достижении максимальной численности быстрее заканчивается летняя репродукция, размножение под снегом не возобновляется; параллельно увеличивается смертность во всех возрастных группах, в итоге численность грызунов резко падает. Год-два после “краха” интенсивность размножения леммингов остается средней, а смертность — сравнительно высокой, и лишь затем популяция вновь вступает в фазу роста. Правильная периодичность пиков и спадов свидетельствует об автономности популяционно-демографических перестроек, формирующих циклы, от факторов среды и о прямой связи их с плотностью популяции.
Растительность
Сравнительно низкая продуктивность тундровых экосистем и огромная численность обитающих там леммингов (в фазе пика до 550 экз. обоих видов на 1 га) позволили предположить, что острая нехватка кормов может стать ведущим фактором в динамике численности грызунов. Действительно, имеются данные о том, что при максимальной численности в некоторых ассоциациях они стравливают растительность на 50—90%. Особенно это характерно для наиболее обедненных тундровых экосистем (подзона арктических тундр) в местах зимней концентрации зверьков.
Благодаря специальным исследованиям связей леммингов и растительности, проведенным на о.Врангеля, мы установили, что сибирские питаются в основном осоками и зелеными мхами, а копытные —кустарничками разнообразных ив и дриад. Злаки и разнотравье оба вида используют в равной мере. Проведенный на модельных куртинах учет фитомассы пяти видов растений (составляющих основу рациона обоих видов леммингов) в год пика и в два последующих года, показал, что осоки и злаки на следующий год после интенсивного стравливания значительно увеличили свою надземную фитомассу. Это говорит о высокой приспособленности тундровой растительности к прессу со стороны леммингов, и в частности о ее способности быстро восстанавливать свою биомассу.
В серии опытов, проведенных на фазе пика, подсчитано, что за три летних месяца копытные лемминги уничтожили от 30 до 55% наземной фитомассы ивнячков и дриад [3]. Поскольку не все побеги и листья кустарничков были доступны грызунам, можно считать, что кормовые запасы в фазе пика используются полностью и появляется дефицит кормов. Наблюдения показали, что зимой, когда площадь пригодных кормов резко сокращается, ситуация осложняется. На следующий год после пика в местах зимовки леммингов растительность была сильно потравлена: в осоково-моховых ассоциациях побеги осок (28% осоки блестящей от общей площади) были целиком объедены. В ассоциациях, где доминировал злак дюпонция Фишера, аналогичные участки занимали 35%.
Сведения, имеющиеся в литературе, и полученные нами данные не дают точного ответа на вопрос, в какой мере дефицит кормов сказывается на выживаемости и репродуктивности леммингов. Тем не менее нехватку кормов, ведущую к нарушению энергетического баланса грызунов, следует рассматривать как вполне реальную причину их зимней смертности, наблюдаемую после пика численности. Но, признавая участие пищевого фактора в регуляции численности леммингов, мы не разделяем точку зрения многих экологов о его ведущей роли.
Хищники
На о.Врангеля, как и в большинстве других районов Арктики, основные потребители леммингов — средний и длиннохвостый поморники (Stercorarius pomarinus, S.longicaudus), белая сова (Nyctea scandiaca) и песец (Alopex lagopus).
В фазе пика наиболее активен средний поморник (он съедает около половины леммингов, уничтожаемых всеми хищниками); в фазе роста (1980) наибольшее количество леммингов приходится на долю белой совы (54.5%), а при постепенном снижении численности — на долю песца (73.5%). Песец, кроме того, единственный, кто охотится на леммингов в фазе их депрессии, когда основные пернатые хищники в тундре не гнездятся.
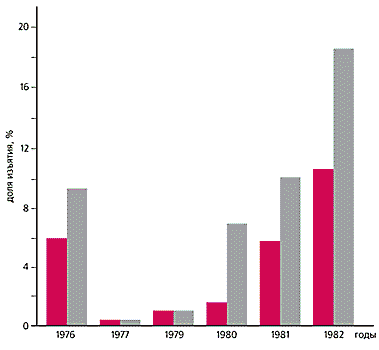
На о.Врангеля нам удалось уточнить реальную величину потерь, которые несут грызуны от хищников в бесснежное время года. Она оказалась в три-четыре раза ниже тех цифр, которые приводятся в работах некоторых авторов по Северной Аляске и Канадскому арктическому архипелагу, районам, где видовой состав хищников почти такой же, как на о.Врангеля. Хотя наши подсчеты не позволяют считать хищников основной причиной падения численности леммингов, видимо, именно они, наряду с факторами демографического характера, сдерживают рост грызунов летом, несмотря на интенсивную репродукцию.
Картина, наблюдаемая на о.Врангеля, характерна лишь для подзоны арктических тундр и, возможно, для северной части типичных тундр. Во многих районах типичных и южных тундр набор пернатых хищников значительно беднее и их пресс явно ослаблен. Именно такая ситуация отмечена нами в Центральной Чукотке и в Чаунской низменности [4], а Ч.Кребс зафиксировал ее в субарктической части Северной Канады [5].
Количество сибирского (выделено цветом) и копытного леммингов, уничтоженных на о.Врангеля в 1976—1982 гг. Наиболее заметен пресс хищников в фазе подъема численности грызунов. Другие внешние факторы
Массовые миграции норвежских леммингов (Lemmus LEMMus) принесли этим зверькам широкую известность задолго до того, как ими заинтересовались ученые. На этот счет существуют многочисленные легенды и предания [1]. Действительно, норвежские лемминги, основные биотопы которых расположены в горных тундрах, в фазе пика во вторую половину лета и осенью сходят с обширных плато, следуя обычно по речным долинам в расположенную ниже лесную зону. Поскольку большинство зверьков при этом погибает, такие миграции естественно резко снижают численность популяции. Как показали наши наблюдения в тундрах северо-востока Сибири и анализ большого количества литературных источников, для сибирского и копытного леммингов характерны сравнительно короткие сезонные перекочевки, которые существенно не меняют общий ход цикла.
В суровых, близких к экстремальным, условиях тундры, где обитают лемминги, климатическая обстановка, казалось бы, могла сказываться на их численности. На это имеется много указаний в литературе. Впрочем, исследования на о.Врангеля говорят о том, что глубина снежного покрова в местах зимнего обитания зверьков, температура воздуха и подснежная температура почти не меняются по годам и, следовательно, заметно не влияют на формирование циклов. То же самое можно сказать и о летних условиях, хотя и более изменчивых по сравнению с зимними.
Во многих, особенно в ранних работах (1942) в качестве одной из основных причин “краха” лемминговых популяций часто упоминались эпизоотии. Действительно, эпизоотии туляремии могут поражать сильно размножившиеся популяции сибирского и копытного леммингов и служить причиной их массовой гибели. Однако природные очаги туляремии обнаружены не во всех районах тундры, к тому же хищники, уничтожающие в основном больных и ослабленных зверьков, сдерживают развитие эпизоотии.
Гипотезы авторегуляции
Традиционно в экологии факторы, влияющие на численность популяции, принято делить на внешние (погода, кормовые ресурсы, хищники, эпизоотии, паразиты, иногда межвидовая конкуренция) и внутренние, которые охватывают весь комплекс внутрипопуляционных взаимодействий, включая физиологические и генетические изменения в ходе цикла, имеющие регуляторное значение. Попытки связать лемминговые циклы с отдельными внешними факторами среды или с их комплексом не имели успеха. В то же время выяснилось, что в ходе циклов у леммингов меняются популяционно-демографические показатели, прежде всего в зависимости от плотности зверьков и лишь потом — от внешних условий (пищи, хищников, погоды, болезней и пр.). Это позволило предположить, что лемминги, подобно многим другим полевкам умеренных широт, обладают адаптивными механизмами авторегуляции. Концепция авторегуляции численности в природных популяциях животных в общем виде разработана В.Вини-Эдвардсом (1962). Применительно к циклическим колебаниям численности мышевидных грызунов эту концепцию развивали в 60—70-х годах Д.Кристиан и Д.Читти.
В целой серии публикаций Кристиана (его первая работа появилась в 1950 г.) подробно изложена гипотеза о возможных нейроэндокринных механизмах регуляции численности мелких грызунов. Хотя в основе этой гипотезы лежала общепризнанная теория Г.Селье о стрессе, она вызвала серьезную критику со стороны некоторых экологов, главным образом из-за недостатка фактических данных о стрессах в природных популяциях [6]. Поэтому исследования, касающиеся роли эндокринного комплекса в регуляции численности грызунов, не получили широкого развития.
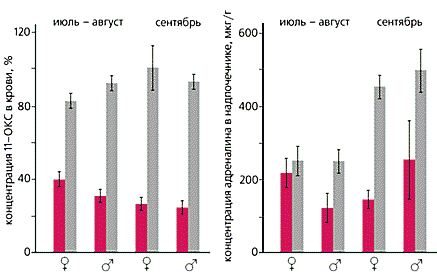
При изучении гормональных перестроек в популяциях леммингов на о.Врангеля установлено, что на разных фазах цикла меняется функционирование не только отдельных желез, но и всего эндокринного комплекса. Наиболее заметно это проявляется в деятельности двух систем: гипофиз—надпочечник, которая наиболее активна в фазе пика численности, и гипофиз—гонады, доминирующая в период депрессии и роста. С динамикой функции надпочечника связана система гипофиз—щитовидная железа. Репродуктивный успех популяции во многом обеспечивается преобладанием адреналового или гонадного фрагментов [3]. Обнаружены изменения не только морфологических (гистологических) показателей ряда эндокринных органов, но и некоторых биохимических параметров, определяющих уровень стресса в популяции [7].
Исследования в Колымской низменности выявили специфические отклонения ряда биохимических и гематологических показателей у сибирского лемминга, свидетельствующие о его повышенном популяционном стрессе в фазе пика [8]. Полученные данные позволяют заключить, что эндокринный комплекс — важная часть авторегуляторных механизмов, участвующих в формировании популяционных циклов леммингов.
В отношении тундровых леммингов в настоящее время имеется немного фактов, которые могли бы подтвердить правильность обсуждаемой гипотезы. Повышенная агрессивность зверьков при максимальной численности хорошо известна зоологам. Характерное для большинства мелких грызунов повышение средней массы в фазе пика и понижение ее в фазе депрессии (так называемый эффект Читти) в полной мере свойственны леммингам. Правильное чередование темпов такого роста в соответствии с фазами цикла можно рассматривать как показатель генетического разнообразия популяции при разной численности.
Таким образом, если роль эндокринного комплекса в регуляции численности леммингов подтверждается фактами, участие генетических перестроек в авторегуляторных реакциях предстоит еще изучить. При этом вполне можно допустить, что оба механизма авторегуляции действуют совместно.
В литературе, посвященной циклам мелких грызунов, появляется все больше работ, ставящих под сомнение реальность гипотезы Читти. В качестве примеров можно привести работу Н.Стенсета (1985) по моделированию циклов, а также эксперименты Р.Бунстры и П.Боага (1987) по выявлению наследуемости некоторых демографических признаков у циклирующей пенсильванской полевки. Да и сам Читти в своей последней книге [10] указывает на существенные трудности, возникающие при попытках экспериментальной проверки своей гипотезы.
Многофакторная модель циклов
Обычно экологи для объяснения феноменологии и механизмов популяционных циклов мелких грызунов используют два методологических подхода: один предусматривает поиск единого механизма, лежащего в основе циклов, другой допускает возможность взаимодействия ряда факторов. Согласно современным представлениям о структуре и функционировании сообществ и экосистем, взаимодействие факторов в динамике численности популяций скорее правило, чем исключение. Значит, численность большинства животных-фитофагов регулируется за счет внутрипопуляционных механизмов и отражает многочисленные биоценотические связи.
Огромный репродуктивный потенциал леммингов (реализуемый, в частности, за счет подснежного размножения), позволяющий им существовать в суровых условиях тундры, обусловливает наличие того или иного механизма контроля над численностью. Без этого достаточно скудный растительный покров тундры мог бы исчезнуть бесследно. Такого, как мы знаем, не происходит. Ряд внешних факторов в фазе пика значительно повышает смертность в популяциях леммингов, а адаптивные механизмы внутрипопуляционной регуляции (эндокринные и, возможно, генетические) воздействуют на все демографические параметры В итоге численность грызунов периодически пульсирует с примерно равными промежутками между пиками. С точки зрения изложенных представлений популяционные циклы — это результат многоуровневых адаптивных реакций и связей, явление, в котором организменные и популяционные механизмы связаны с процессами, происходящими в биоценозе.
Работа выполнена при поддержке Российского фонда фундаментальных исследований.
Проекты 93-04-07962 и 02-04-48024.Литература
1. Чернявский Ф.Б. Загадка лемминговых циклов // Природа. 1968. №12. С.45—50.
2. Stenseth N., Ims В. (Ed.) The biology of lemmings // Lin. Soc. Symp. Acad. Press. 1993. №15.
3. Чернявский Ф.Б., Ткачев А.В. Популяционные циклы леммингов в Арктике. Экологические и эндокринные аспекты. М., 1982.
4. Чернявский Ф.Б. и др. // Фауна и экология млекопитающих северо-востока Сибири. Магадан, 1995. С.82—92.
5. Krebs С J. // Techn. Paper Arct. Inst. North Amer. 1964. №15.
6. Lee А.К., McDonald J.К. // Oxford Rev. Reprod. Biol. 1985. V.7. P.261—304.
7. Чернявский Ф.Б., Дорогой И.В. и др. // Доклады АН СССР. 1978. Т.242. №3. С.730—733.
8. Чернявский Ф.Б., Лазуткин А.Н. // Сиб. экол. журн. 1999. №1. С.99—105.
9. Chitty D. // Canad. J. Zool. 1960. V.38. №1. P.99—113.
10. Chitty D. Do Lemmings Commit Suicidal? Beautiful Hypothesis and Ugly facts. Oxford, 1996.