№11, 2003 г.
© В.П. НикишинНеобычная организация
покровной ткани скребнейВ.П. Никишин
В обширном и разнообразном мире паразитических организмов несколько особняком стоят те, которых в обиходе уже давно называют паразитическими червями, или гельминтами. Группа эта сборная, включает животных нескольких типов, весьма отличающихся по морфологии и экологии. В большинство из них входят как паразитические, так и свободноживущие виды (коловратки, плоские и круглые черви), и только тип скребней (Acanthocephala) представлен исключительно паразитирующими организмами (рис.1). По сравнению с другими гельминтами скребни изучены довольно слабо, и это понятно - их видовое разнообразие невелико, да и вызываемые ими акантоцефалезы довольно редкие заболевания животных и тем более человека.Владимир Павлович Никишин, доктор биологических наук, ведущий научный сотрудник
лаборатории популяционной генетики Института биологических проблем Севера Дальневосточного отделения РАН.
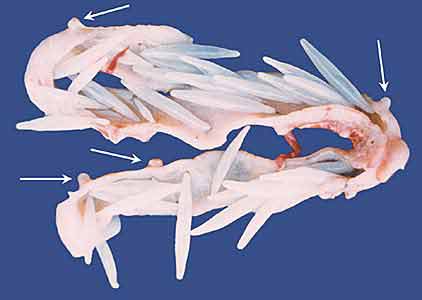
Рис.1. Вскрытый участок кишечника утки-шилохвостки (Anas acuta), пораженный зрелыми самками скребней (Filicollis anatis). Хоботки скребней глубоко внедряются в стенку кишечника и затем разрастаются, образуя на внешней поверхности кишки выросты в виде луковицы (показано стрелками). Фото Г.И.АтрашкевичаВо взрослом состоянии скребни заселяют только кишечник, обычно тонкую кишку, у всех групп позвоночных животных (у рыб - еще и пилорические отростки). Иногда они, пробуравливая стенку кишечника, проникают в полость тела, но это уже явная патология как для хозяина, так и для самого скребня, который никогда не сможет оставить потомства. Личинки развиваются в ракообразных и насекомых (рис.2), причем с весьма высокой специфичностью: среди нескольких близкородственных видов промежуточных хозяев тем или иным видом скребней заражается только один. Так, промежуточным хозяином скребня Moniliformis moniliformis siciliensis может быть только жук чернотелка, M.m.moniliformis - еще и американский таракан, Macracanthorhynchus hirudinaceus - майский жук. Понятно, почему люди так редко страдают акантоцефалезами, ведь для этого по крайней мере надо проглотить таракана или жука.
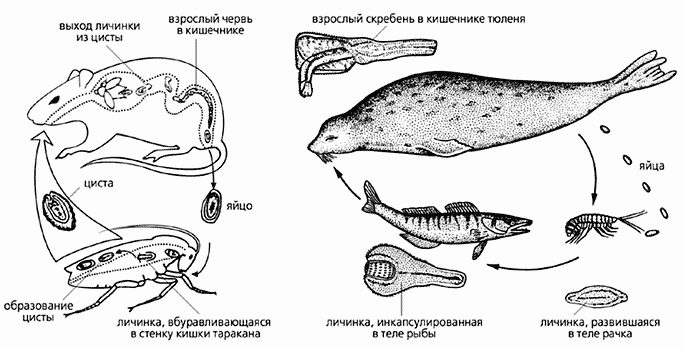
Рис.2. Жизненные циклы двух видов скребней.
Один (из рода Moniliformis) паразитирует в кишечнике грызунов (слева), другой (Corynosoma strumosum) - тюленей.Морфология скребней на первый взгляд довольно однообразна. Обычно это небольшие черви длиной до одного-двух сантиметров, и лишь некоторые виды достигают больших размеров. Например, скребень-великан M.hirudinaceus (паразит свиньи, а иногда и человека) вырастает до полуметра и более. Ланцетовидное тело скребней отчетливо делится на две части (рис.3). Передняя (пресома) - состоит из относительно большого и подвижного хоботка, вооруженного крючьями (их количество и расположение служат основными таксономическими признаками), а также из небольшой шейки. В задней части (метасоме) размещены органы половой системы и выделительной (в случае ее наличия). Пищеварительный тракт отсутствует. Скребни осмотрофы, т.е. питаются, всасывая необходимые вещества всей поверхностью тела. Своеобразно устроен и эпителий - не из отдельных клеток, как у большинства многоклеточных животных, а в виде сплошного цитоплазматического пласта, в котором ядра (кстати, их число ограничено, что также служит систематическим признаком), как и у обычных эпителиальных клеток, располагаются в базальной части. Получается, что весь покров скребней образован единой многоядерной клеткой (симпластом), которой свойственны функции не только “нормального” эпителия, входящего в состав кожного покрова (защита, опора, транспорт, секреция и т.д.), но и пищеварительных эпителиев (всасывание, секреция ферментов и т.д.). Чтобы подчеркнуть столь необычную организацию и многофункциональность пограничной ткани скребней, для нее был принят специальный термин “тегумент” [1] (см. рис.3).
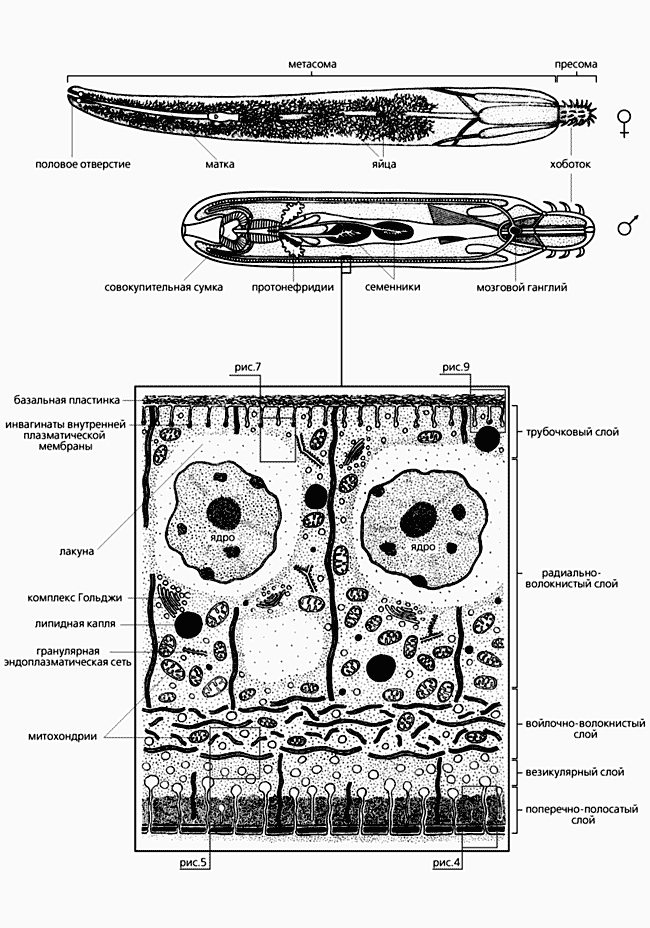
Рис.3. Схема строения тела скребней и отдельно тегумента метасомы.
У других паразитических червей, а именно у сосальщиков (Trematoda) и ленточных червей (Cestoda), пограничная ткань также представлена тегументом, но организованным совсем иначе. У этих гельминтов только дистальные части эпителиальных клеток объединены в сплошной пласт, а базальные, содержащие ядра и большую часть органелл и включений, погружены глубоко под слои субповерхностных мышц и не часто (обычно на стадиях интенсивного роста организма) связаны между собой цитоплазматическими мостиками. Конструкции, когда клетки связаны друг с другом фрагментарно, а их ядросодержащие части представляют собой отдельные “карманы”, аналогичные клеточным телам, называются синцитиями. Таким образом, тегументом обозначают две совершенно разные по организации пограничные ткани: у скребней это симпласт, а у плоских червей - синцитий. Кстати, впервые термин “тегумент” был введен при исследовании покровной ткани именно ленточных червей.
Зачем же понадобилось объединять эпителиальные клетки в единое целое столь кардинальным способом, и чем не устраивала паразитических червей клеточная конструкция эпителия? Разумеется, возникновение тегумента было напрямую связано с паразитическим образом жизни его обладателей, а точнее с необычными условиями среды обитания. Не так уж важно в этом случае, имели ли предки ныне живущих цестод, трематод или скребней пищеварительный тракт или нет, поскольку освоение новой среды открыло принципиально новые возможности питания. Вместо формирования громоздкого пищеварительного аппарата и развития инструментов, необходимых для поиска и захвата пищи, в новых условиях существования можно было просто воспользоваться пищеварительной системой хозяина и всасывать уже готовые или почти готовые продукты.
Другим необходимым условием возникновения тегумента было отсутствие у низших многоклеточных, к числу которых относятся предки нынешних гельминтов, сосудистых систем, доставляющих необходимые вещества к разным участкам тела. Конечно, существует немало механизмов переноса тех или иных субстанций сквозь клеточную стенку (цитоплазматическую мембрану), однако действуют они довольно медленно и в большинстве случаев лишь гармонично дополняют работу сосудистых систем. Частичная утрата цитоплазматических мембран и превращение клеточного эпителия в симпластический или синцитиальный, естественно, облегчает задачу, поскольку для транспорта используются внутриклеточные системы (микротрубочки, эндоплазматический ретикулум и т.д.). Более того, у скребней, например, в толще тегумента развита система своеобразных каналов - лакун.
И наконец, отказаться от клеточного строения эпителия гельминтов вынудила весьма агрессивная среда обитания - организм хозяина, чутко реагирующий на вторжение, будь то паразит или обычная заноза. И если в отношении питания или транспорта питательных веществ образ жизни привел к определенному упрощению организации паразитических червей, то для защиты от агрессивного ответа хозяина на вторжение у них выработались весьма совершенные и разнообразные механизмы. Несомненно, один из них - возникновение тегумента, благодаря чему гельминты получили возможность оперативно и адекватно отвечать на нежелательные для паразита реакции хозяина всей поверхностью тела.
Справедливости ради замечу, что не у всех паразитических червей покровная ткань трансформировалась в тегумент. У многих нематод (Nematoda) эпидермис образован отдельными, правда, очень немногочисленными клетками [2]. Но и у некоторых представителей этого класса в процессе личиночного развития изначальный клеточный эпидермис становится синцитиальным. И это при том, что все нематоды защищены мощной и сложно устроенной кутикулой, которая образуется в результате секреции ее материала эпителиальными клетками (или синцитием). У плоских червей (к коим относятся трематоды и цестоды) и у скребней такой кутикулы нет.
Тегумент плоских червей иногда называют погруженным эпителием, чем подчеркивают одну из основных его особенностей - расположение клеточных тел под слоями кожной мускулатуры. Биологический смысл явления очевиден: в этом случае наиболее важная часть клеток, содержащая ядра и основную долю клеточных органелл и включений, надежно защищена от механических воздействий организма хозяина поверхностным слоем цитоплазмы, базальной пластинкой и мышечными слоями. Считается, что такая конструкция в эволюционном отношении более прогрессивна, чем, скажем, клеточный эпидермис низших турбеллярий, располагающийся над базальной пластинкой и слоями кожной мускулатуры. Нечто похожее наблюдается и у нематод, у которых основные порции цитоплазмы эпидермальных клеток вместе с большей частью клеточных органелл, в том числе и ядрами, располагаются в так называемых гиподермальных валиках, погруженных глубоко под кожную мускулатуру.
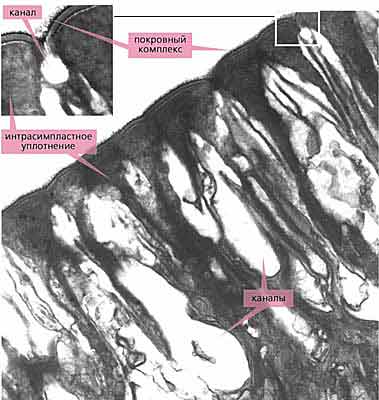
У скребней ничего подобного нет, и их симпластический тегумент покоится над базальной пластинкой и слоями кожной мускулатуры. Однако для защиты от механических повреждений у них развились достаточно эффективные и отсутствующие у других паразитических червей структуры. Во-первых, это покровный комплекс из поверхностной цитоплазматической мембраны и расположенных непосредственно под ней одной-двух плотных пластинок, который формируется еще на стадии эмбриона и в дальнейшем не меняется (рис.4).
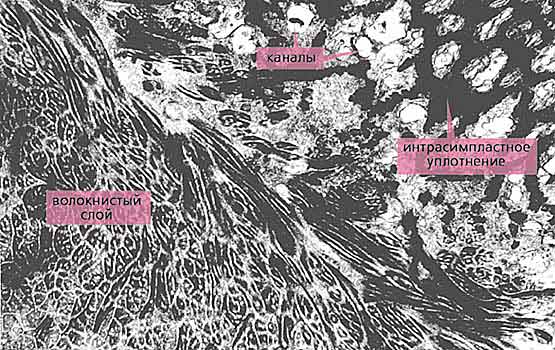
Вплотную к внутренней стороне покровного комплекса примыкает вторая, наиболее укрепленная защитная система - интрасимпластное уплотнение - мощный (до нескольких микрометров толщиной) слой плотного материала [3]. Прежде, а иногда и сейчас, интрасимпластное уплотнение ошибочно считали кутикулой. На самом деле это не так, поскольку располагается оно не на поверхности, а внутри эпидермиса (покровного симпласта), формируется, когда скребень попадает в организм окончательного хозяина, и не подвергается при линьке. Наконец, еще глубже находится самая объемная, третья линия обороны - система горизонтальных волокон, ориентированных в продольном или кольцевом направлении (рис.5).
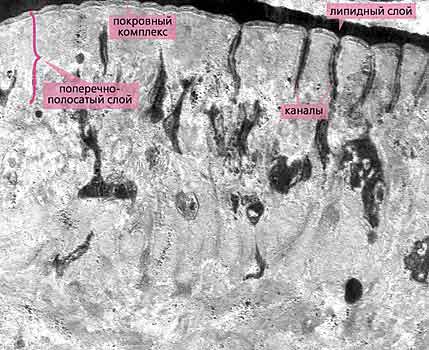
Рис.5. Дистальная часть войлочно-волокнистого слоя тегумента метасомы скребня Polymorphys magnus. Заметно упорядоченное расположение волокон, каждое из которых состоит из нескольких субъединиц. В поперечно-полосатом слое видны расширенные основания каналов, между которыми находится интрасимпластное уплотнение. Увел. 12 800.Эти волокна образуют войлочно-волокнистый (или просто войлочный) слой тегумента, степень развития которого зависит от окончательного хозяина. Например, у рыбьих скребней эти волокна менее многочисленны и неупорядочены, поскольку пищеварительный тракт рыб оказывает на паразитов меньшие механические воздействия, чем, скажем, кишечник околоводных птиц. Волокна войлочного слоя у птичьих скребней упорядочены и организованы в несколько (до пяти) субслоев. Разумеется, все эти образования выполняют не только защитную функцию, но и опорную [4], за которую у многих других беспозвоночных ответственна кутикула.Казалось бы, столь действенные защитные системы должны препятствовать движению всасываемой пищи в глубь тегумента для дальнейшей переработки. Без сомнения, они и препятствуют, и поэтому эволюция снабдила скребней великолепным механизмом, позволяющим не только преодолеть ею же созданные барьеры, но и многократно увеличить всасывающую поверхность.
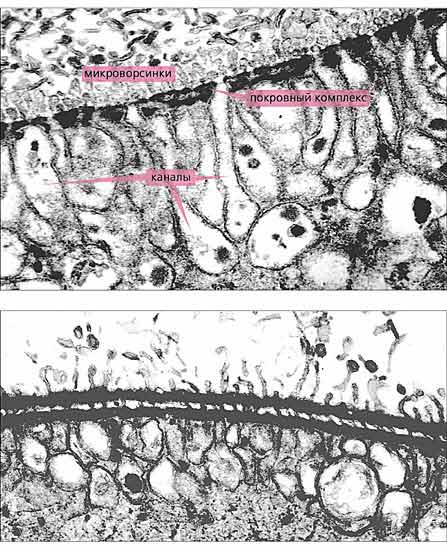
У цестод, как известно, на поверхности тегумента формируются специализированные микроворсинки (микротрихии), выполняющие помимо всасывания и другие функции. Но у цестод нет покровного комплекса и интрасимпластного уплотнения, поэтому такая конструкция их вполне устраивает. У скребней же весь поверхностный отдел тегумента пронизан каналами, открывающимися на поверхности червя и слепо заканчивающимися в глубине тегумента. (Благодаря этим каналам поверхностный отдел тегумента назван поперечно-полосатым, или просто полосатым, слоем). Причем заканчиваются каналы значительно глубже интрасимпластного уплотнения и обязательно в виде расширенных пузырьков. Стенки каналов и соответственно концевых пузырьков образованы только цитоплазматической мембраной и не имеют покровного комплекса, поэтому ничто не мешает перемещению всасываемых субстанций в глубь тегумента. Что же касается площади поверхности цитоплазматической мембраны, через которую осуществляется питание червей, то, по разным оценкам, оно может достигать 20-, 40- и даже 60-кратного увеличения в зависимости от степени ветвления каналов, а также вида и экологии скребня. Надо сказать, что функции каналов не ограничиваются питанием, но об этом чуть позже.
На поверхности тегумента у скребней есть и микроворсинки, но очень короткие, и развиваются они только в период личиночного развития, когда червь находится в организме промежуточного хозяина (рис.6). (В этот период в тегументе еще нет интрасимпластного уплотнения, однако система каналов в поверхностном отделе тегумента, а также покровный комплекс уже сформировались.) Вершинки микроворсинок постепенно отделяются и входят в состав окружающей личинку цисты. Незадолго до завершения личиночного развития микроворсинки уже целиком отделяются от тегумента и в течение дальнейшей жизни скребней на поверхности тегумента больше не развиваются [5].
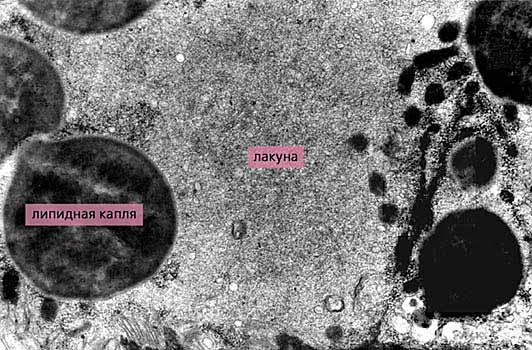
Рис.6. Тегумент личинок Filicollis anatis разного возраста. У 50-дневных личинок (вверху) многочисленные микроворсинки имеют “нормальную” внешность, а у 60-дневных они уплотнены и служат своеобразной арматурой для слоя плотного материала - будущей внутренней части цисты. Увел. 21 500.Оригинальной особенностью тегумента скребней является не имеющая или почти не имеющая аналогов в животном мире транспортная система, о которой уже вскользь упоминалось. Речь идет о лакунах - системе каналов, сосредоточенных в базальной половине тегумента и пронизывающих его вдоль и поперек. Строго говоря, это вовсе и не каналы, поскольку не имеют собственных стенок. На электронных микрофотографиях они выглядят как участки цитоплазмы, полностью лишенные органоидов и лишь изредка содержащие немногочисленные включения, главным образом липидные капли (рис.7). До сих пор не ясно, как лакуны сохраняют свою форму и почему органоиды из окружающей цитоплазмы не перемещаются в них?
Рис.7. Фрагмент лакуны, окруженный липидными каплями, митохондриями и волокнами, из внутренней части тегумента метасомы зрелой личинки скребня Polymorphys magnus. Увел. 15 400.Сказанным особенности лакун не исчерпываются. У скребней тегументы метасомы и пресомы на уровне основания шейки разделены особой перегородкой, и в тегументе каждого отдела есть своя лакунная система. Кроме того, введением различных веществ, в том числе и красящих, обнаружена еще одна лакунная система, на этот раз в субповерхностных мышцах, которая сообщается с лакунной системой тегумента метасомы [6]. Это означает, что содержимое лакун может перемещаться при сокращении и расслаблении мышц. Кстати, кожная мускулатура скребней в функционально-морфологическом отношении заслуживает не меньшего внимания, чем тегумент, но это уже другая тема.Как упоминалось, в число обязательных функций тегумента входит выделение, или секреция. Об одном из проявлений этой функции - отделении вершин микроворсинок у личинок скребней (что в сущности не что иное, как секреция по микроапокринному типу) - уже говорилось. Подобное явление достаточно часто встречается в природе, и характерно, например для ленточных червей. Однако тегументу скребней присущи еще, как минимум, два типа секреции, не отмеченных у других животных, в том числе и паразитических. Оба они осуществляются с участием каналов поперечно-полосатого слоя. Один из них (тоже упомянутый) наблюдается в конце личиночной стадии, когда разово и быстро выделяется плотный материал (его состав пока не исследован), образующий на поверхности личинки сплошной слой, пронизанный микроворсинками. Впоследствии этот слой вместе с микроворсинками отделяется от тегумента и формирует внутреннюю часть цисты [5] (см. рис.6).
Секреция другого типа - выделение липидной субстанции, образующей на поверхности тегумента мощный электронно-плотный слой; происходит это в пресоме у большинства взрослых скребней, паразитирующих в окончательном хозяине [7] (рис.8). Возможно, липидный слой служит своеобразной смазкой, которая облегчает скребню внедрение в ткани хозяина, или защищает паразита от отторжения.
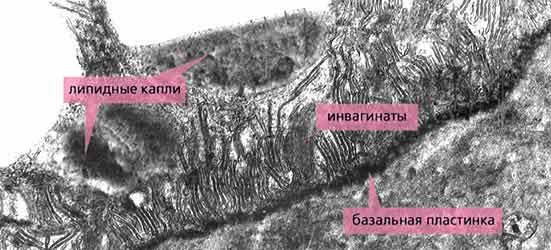
Рис.8. Тегумент пресомы скребня Polymorphys magnus. Поверхность тегумента образована покровным комплексом, под которым располагается поперечно-полосатый слой, пронизанный каналами. Интрасимпластное уплотнение заметно слабо. Каналы заполнены тем же плотным материалом, из которого образован липидный слой на поверхности. Увел. 28 000.Но и это еще не все. Например, без секреции тегументом пресомы ферментов, разрушающих ткани хозяина, червям было бы трудно или даже невозможно закрепиться в каком-либо участке кишечника. Тегумент метасомы также выделяет на свою поверхность ферменты, но уже другие, участвующие в переваривании пищи. Однако эти и некоторые другие виды секреции уже не столь уникальны, поскольку встречаются у других паразитических червей.Уникален тегумент скребней и по сложности строения. Помимо уже упомянутых слоев - покровного комплекса, поперечно-полосатого (пронизанного каналами интрасимпластного уплотнения), везикулярного (состоящего из расширенных окончаний каналов), войлочного (образованного многочисленными волокнами), радиально-волокнистого (своеобразного метаболического центра, в котором перерабатываются поглощенные поверхностью субстанции и откладываются запасные вещества) - есть еще так называемый трубочковый слой (см. рис.3). Он образован многочисленными выростами внутренней цитоплазматической мембраны тегумента, направленными в его толщу и имеющими внешность трубочек (рис.9), благодаря которым он и получил свое название [8]. Вообще подобная организация свойственна всем или почти всем эпителиальным клеткам, а также поверхностным синцитиям других паразитических червей. Однако у скребней элементы этого слоя наиболее многочисленны, а сам слой имеет наибольшую толщину по сравнению с другими эпителиями. Его функциональное значение, как и у других эпителиев, заключается в обмене материалами между тегументом и “полостью” тела, в том числе в транспорте воды и ионов [9].
Рис.9. Трубочковый слой тегумента метасомы зрелой личинки скребня Polymorphys magnus. Инвагинаты внутренней цитоплазматической мембраны тегумента - радиально ориентированные трубочки - расположены очень плотно друг к другу. В прилегающей цитоплазме видны липидные капли. Увел. 21 000.* * * В многообразном царстве животных есть лишь одна группа организмов, покровная ткань которых во многом сходна с тегументом скребней, - коловратки (Rotatoria). Это мелкие (от 0.04 до 2 мм), в основном свободноживущие (пресноводные) черви, обладающие пищеварительным трактом (т.е. питающиеся “нормальным” способом) и реснитчатым коловращательным аппаратом для движения. Удивительно, но именно коловратки имеют симпластический эпителий, аналогичный (часто до мельчайших деталей) тегументу скребней - паразитирующих организмов. Что это? Конвергенция? Маловероятно. Филогенетическая связь? Вполне возможно. Сходство с общим предком? Очень может быть. Среди коловраток ведь есть внутренние (кишечные) паразиты беспозвоночных (преимущественно малощетинковых червей). В результате паразитирования некоторые коловратки даже утратили коловращательный аппарат. Возможно, именно от таких форм и берут начало отдаленные предки скребней. Тем более что сходство в строении покровов коловраток и скребней дополняется общим планом строения нервной системы (мозговой ганглий и одна пара боковых нервных стволов) и наличием клоаки, куда у скребней выходят отверстия половой и выделительной систем, а у коловраток еще и пищеварительной. Кроме того, симпластический тегумент не менее логично выводится из эпителия турбеллярий, чем синцитиальный - из эпителия плоских паразитических червей. По крайней мере для обеих разновидностей тегумента найдены промежуточные формы эпителия, присущие современным турбелляриям.А может быть, обе группы (коловратки и скребни), учитывая их предполагаемую (и на то есть основания) древность, представляют собой остатки некогда весьма прогрессивного направления в эволюции беспозвоночных?
В любом случае загадочное сходство покровов этих, столь различных по образу жизни, животных еще ждет своего изучения. А пока остается констатировать, что еще одной загадкой в мире живой природы стало больше.
Разрешима ли она?
Работа поддержана Президиумом Дальневосточного отделения РАН (конкурс 2002 г.). Проекты А-36 и К-8.
Литература
1. Miller D.M., Dunagan T.T. Functional morphology // Biology of the Acanthocephala. Cambridge, 1985.
2. Малахов В.В. Нематоды: строение, развитие, система и филогения. М., 1986.
3. Вельш У., Шторх Ф. Введение в цитологию и гистологию животных. М., 1976.
4. Богоявленский Ю.К., Иванова Г.И. Микроструктура тканей скребней (покровы и нервная система). М., 1978.
5. Nikishin V.P. // J. Parasitol. 1992. V.78. №1. P.127-137.
6. Miller D.M., Dunagan T.T. // Proc. Helminthol. Soc. Wash. 1985. V.52. №2. P.221-226.
7. Taraschewski H. // Adv. Parasitol. 2000. V.46. P.1-179.
8. Никишин В.П. // Паразитология. 2000. Т.24. №2. С.125-143.
9. Куперман Б.И. Функциональная морфология низших цестод: онтогенетический и эволюционный аспекты. Л., 1988.