№11, 2004 г.
№11, 2004 г.
© Шарова Н.П., Абрамова Е.Б.
Повреждение и починка ДНК,
или
“На всякую прореху найдется заплата”Н.П.Шарова, Е.Б.Абрамова
Наталья Петровна Шарова, доктор биологических наук, ведущий научный сотрудник Института биологии развития им.Н.К.Кольцова РАН.
Елена Борисовна Абрамова, кандидат биологических наук, старший научный сотрудник того же института.Записанная в ДНК генетическая программа любого живого существа - от простейшего одноклеточного до высокоорганизованного многоклеточного - сохраняется в череде потомков благодаря точному воспроизведению нуклеотидной последовательности в каждом поколении. Но если в главной наследственной молекуле возникают повреждения, а это случается даже спонтанно, изменения в ее структуре могут привести не только к разным патологиям тканей или органов, но и к передаче мутации потомству. И, как это ни удивительно, ДНК - единственная клеточная макромолекула, способная исправлять повреждения в собственной структуре. В ней даже закодирована информация о механизмах тех процессов, которые занимаются таким ремонтом (в строго научной среде их называют репарационными). Именно “починка” ДНК обеспечивает запас прочности организма.
Восстановление структуры ДНК особенно важно при формировании репродуктивных (половых) клеток и в эмбриональном развитии: во время активного синтеза новых цепей риск появления и повторения ошибок многократно увеличивается. Наследственная молекула должна залечиваться очень быстро, и делящиеся эмбриональные клетки “знают”, как это осуществить. Устраняются разные повреждения ДНК и в течение всей жизни организма. Начнем рассказ с известных к настоящему времени типов повреждений, чтобы затем рассмотреть механизмы починки, потому что от характера неисправности зависит, каким способом она будет ликвидирована.
Что может испортиться в ДНК
Поломки возможны в любом из образующих ДНК компонентов - и в азотистых основаниях, и в сахарофосфатном остове, причем как при копировании (репликации), так и при считывании (транскрипции) информации для последующего синтеза клеточных белков.
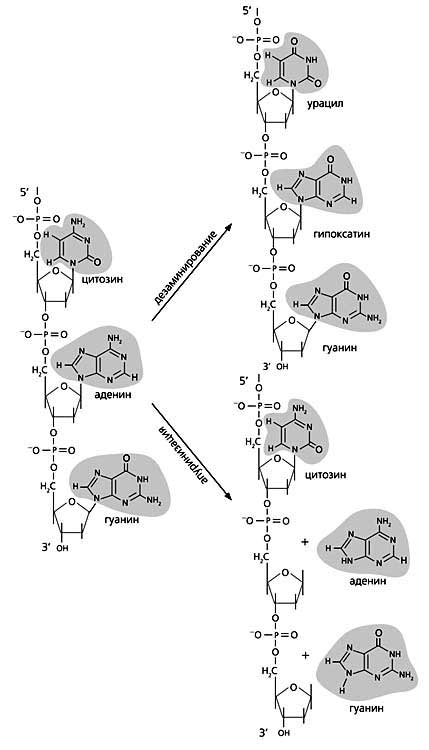
Часто случаются апуринизация того или иного нуклеотида и дезаминирование оснований. В первом случае рвутся гликозидные связи между пурином (аденином или гуанином) и дезоксирибозой, в результате чего эти основания выщепляются из цепи ДНК (место, в котором произошло такое событие, называют АП-сайтом). Второй случай - дезаминирование - приводит к образованию несвойственных для структуры ДНК соединений: вместо цитозина, аденина или гуанина появляются урацил, гипоксантин или ксантин, соответственно. Оба процесса спонтанные. За сутки в клетке человека апуринизация повторяется 5-10 тыс. раз, а частота дезаминирования составляет примерно 100 событий на полный геном.
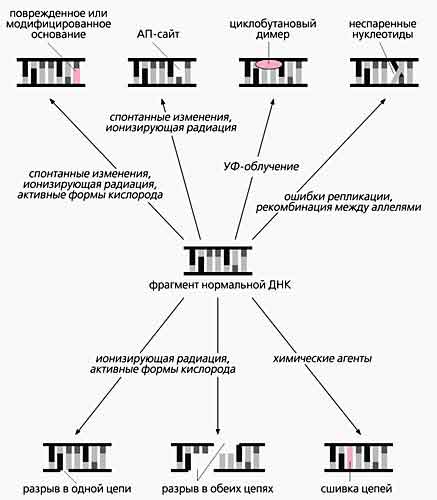
Действие ультрафиолетового облучения приводит к насыщению двойных связей пиримидиновых оснований и образованию димеров из двух соседних пиримидинов в одной цепи ДНК. Ионизирующая радиация может вызвать несколько повреждений: разрыв пуринового кольца, фрагментацию основания, окисление апуринового сайта, а также одно- и двухцепочечные разрывы (это фактически разлом хромосом - главная причина летального действия ионизирующей радиации). Некоторые химические агенты способны сшивать цепи ДНК. Активные формы кислорода (ОН·, О2·–, Н2О2, перекиси липидов и др.), постоянно генерируемые в процессах метаболизма, повреждают и основания, и дезоксирибозу, что способствует образованию новых ковалентных связей. Особенно подвержены окислительному действию соседствующие гуанины (GG). Это буквально “горячие точки” такого процесса, а его конечный результат - модифицированное производное гуанозина. Сахарофосфатные связи разрушаются в тех случаях, когда обе нити ДНК лишились пуринов в противолежащих друг другу местах или вблизи произошла фрагментация дезоксирибозы. Тогда рвутся сразу обе цепи ДНК. Источником структурных дефектов может быть и естественный процесс - репликация, если в комплементарных цепях появляются неспаренные нуклеотиды.
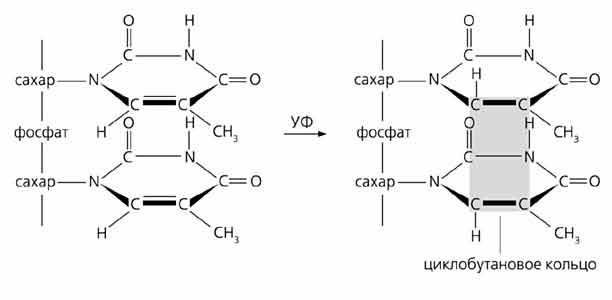
Слева - Нормальная молекула ДНК (фрагмент) и схемы образования наиболее частых повреждений - дезаминирования, апуринизации и циклобутановых димеров. При дезаминировании цитозин превращается в урацил, а аденин - в гипоксантин. Если разрывается связь между остатком дезоксирибозы и пуриновым основанием (здесь аденин и гуанин), в этом месте остается только сахарофосфатный остов, т.е. возникает апуриновый сайт. Под действием ультрафиолета в ДНК может образоваться циклобутановый димер из двух соседних пиримидинов (тиминов), находящихся в одной цепи (вверху).Внизу - Повреждения, которые могут возникать в ДНК спонтанно или под действием разных агентов.
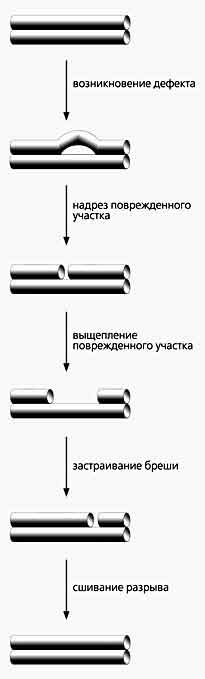
Клетка способна устранить перечисленные повреждения, несмотря на их различия, и восстановить структуру ДНК. Вполне понятно, что для этого требуются специальные механизмы. И хотя в частностях они отличаются один от другого и бывают весьма сложными, все же подчиняются общим принципам. Сначала опознается вид возникшей неисправности. Этим занимаются один белок или несколько (тогда они объединяются в месте изъяна в комплекс). Затем поврежденный участок вырезается в ходе ферментативных реакций, после чего ДНК-полимераза синтезирует правильный кусок ДНК. Завершается репарация сшиванием отдельных фрагментов цепи ДНК-лигазой. Такова общая схема, но каждый тип ремонта осуществляют собственные белки и ферменты, определяющие его индивидуальность. Не правда ли, эта схема напоминает обычное латание дыр на одежде?
Молекулярные ножницы и заплатки
Большая часть структурных нарушений в ДНК устраняется вырезанием (этот тип репарации специалисты называют эксцизионной; по-англ. excision repair), которое осуществляют ферменты нуклеазы. Бракованное основание удаляется в одиночку или вместе с окружением - участками цепи в 2-10 нуклеотидов. Так вырезаются модифицированные и поврежденные основания, апуриновые сайты и места с одноцепочечными разрывами. (Данная разновидность названа эксцизионной репарацией основания; base excision repair, BER.) В случае более крупных неисправностей - возникновения пиримидиновых димеров или других объемных образований в ДНК, нарушающих структуру спирали, - клеточные ножницы отстригают испорченную часть в составе куска длиной в 30 нуклеотидов. (Это уже эксцизионная репарация нуклеотида, nucleotide excision repair, NER.)
Обобщенная схема восстановления нормальной структуры ДНК. Дефектным может быть одиночное основание, и тогда вырезается только нуклеотид; в случае более объемного повреждения из цепи удаляются и соседние с ним нуклеотиды, а образовавшуюся брешь застраивают полимеразы - или самостоятельно, или в присутствии белка-помощника (PCNA). Каков будет размер вырезаемого куска, а значит, и заплаты, зависит от самого повреждения и соотношения имеющихся в клетке ДНК-полимераз. Эти ферменты застраивают бреши, образовавшиеся после вырезания аномального участка, используя в качестве матрицы фрагмент противоположной цепи ДНК. ДНК-полимеразы (у эукариот их известно более 15) различаются по ряду признаков, в том числе по количеству нуклеотидов, которые они успевают встроить в растущую цепь за один акт связывания с дуплексом ДНК. Полимеразы b и l присоединяют всего один нуклеотид, т.е. накладывают маленькую заплатку (short patch путь BER, как говорят специалисты). Две другие ДНК-полимеразы - d и e - способны создать большую вставку, но для этого им нужен специальный помощник - белок PCNA (proliferating cell nuclear antigen). Особенно зависим от него первый фермент - без помощника он “отваливается” от ДНК, встроив лишь один нуклеотид, а если PCNA удерживает на ней полимеразу d, то синтез идет до тех пор, пока фрагмент не достигнет нужной длины. Если в клетке не хватает той или иной ДНК-полимеразы, репарация может переключиться с одного пути на другой, т.е. с малой заплаты на большую (long patch путь BER). Клеточные линии фибробластов мышиных эмбрионов восстанавливают нормальную структуру на месте апуринизации преимущественно посредством малой заплатки (в 80% событий), а если в клетках поврежден ген ДНК-полимеразы b, починка проходит исключительно наложением большой заплаты. Кстати, способ латания дыр клеткой зависит и от того, сколько в ней содержится белка PCNA.
Мелкие неполадки. Если в ДНК изменено одно основание, его распознают, а потом вырезают N-гликозилазы, и в результате в цепи остается дезоксирибозофосфат (как при апуринизации). Далее сахарофосфатный остов надрезается АП-эндонуклеазой (класса II) с 5ў-стороны от этого промежуточного продукта, похожего на АП-сайт. Совсем удалить его клетка может двумя способами (short patch или long patch BER): вырезав только сам остаток дезоксирибозофосфата или вместе с окружающими двумя-десятью нуклеотидами. По размеру образовавшейся дырки синтезируется и заплата. Брешь в один нуклеотид застраивается ДНК-полимеразой b. Иное дело, если появившийся в ДНК АП-сайт еще и подвергся окислению или восстановлению. Такую структурную аномалию вместе с окружающими нуклеотидами вырезает другой фермент - нуклеаза FEN1, - а образовавшуюся брешь застраивают ДНК-полимеразы d или e, которые работают только с помощником - белком PCNA. Иногда короткие пробелы (длиной до 5-6 нуклеотидов) заполняют ДНК-полимеразы b или l. Помощник им не нужен, так как они сами могут временно “заякориться” на свободном 5ў-конце цепи, прилежащем к бреши, и задержаться на какое-то время на ДНК, чтобы осуществить ресинтез вырезанного участка.
Мы несколько лет занимаемся исследованием репарации у костистых рыб в период их эмбрионального развития. В этот период онтогенеза активно синтезируются новые цепи ДНК, и потому риск появления и повторения ошибок многократно увеличивается. Чтобы не произошло подобного, наследственная молекула должна очень быстро залечиваться. Сначала мы предположили, что делящиеся эмбриональные клетки должны иметь несколько равноценных способов устранения одного и того же повреждения. Таким способом одновременно восстанавливалась бы структура ДНК на всех поврежденных участках, и процесс успешно завершался бы даже в том случае, если другие пути репарации подавлены по какой-либо причине. Но вместо того, чтобы экспериментально подтвердить свое предположение, мы обнаружили неизвестный ранее вид репарации ДНК. Работая с эмбрионами и яйцеклетками вьюна (костистой рыбы), мы неожиданно столкнулись с самостоятельной застройкой довольно крупных брешей (до 10-13 нуклеотидов) ДНК-полимеразой d - тем самым ферментом, который, как только что упоминалось, не может обходиться без помощника PCNA. Но в эмбриональном периоде вьюна полимераза d в помощнике не нуждалась. Оказалось, что этот фермент способен связываться одновременно с обоими концами разорванной ДНК [1] и прочно удерживаться там, пока не “залатает дырку” полностью. Этот механизм репарации ДНК в эмбриональных активно делящихся клетках ранее не был известен. Между тем он очень важен, особенно если учесть, что большой заплатой устраняется основное число возникших в ДНК дефектов. Открытый нами способ репарации уникален тем, что действует в экстремальных условиях: совсем без белка-помощника или при его незначительном количестве, а также в том случае, когда в клетке не хватает других ДНК-полимераз, которые обходятся без PCNA.
Не дырка, а затяжка. Ультрафиолетовые лучи вызывают ковалентное сшивание соседних пиримидинов (например, тиминов) в молекуле ДНК. В распознавании и вырезании образовавшихся циклобутановых димеров участвуют семь белков - продуктов генов семейства ХР (XPA-XPG). У человека мутации хотя бы в одном из них могут привести к возникновению наследственного заболевания - пигментной ксеродермы (xeroderma pigmentosum, ХР), которая проявляется в первые годы жизни ребенка. На не защищенных от солнечных лучей участках кожи появляются пигментные пятна и в конце концов может развиться рак кожи.
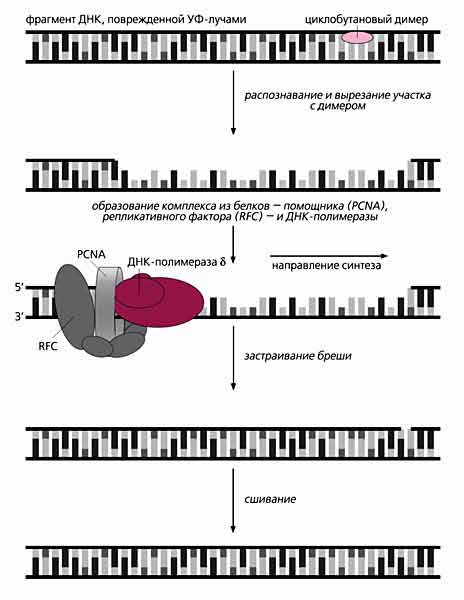
Зачем нужно так много белков, и как они действуют? Каждый из них играет строго определенную роль. Белок ХРА узнает пиримидиновый димер в ДНК и связывается с другими членами семейства. ХРВ и XPD входят в состав сложного белкового комплекса - фактора транскрипции IIH (TFIIH), который расплетает ДНК в участке, подлежащем вырезанию. Еще один белок - ХРС - удерживает этот комплекс на поврежденном участке ДНК, а два последних (их называют нуклеазами вырезания) - XPF и XPG - работают как ножницы. Первая нуклеаза разрезает 24-ю фосфодиэфирную связь с 5ў-стороны от димера, а вторая - 5-ю связь с 3ў-стороны. В результате их совместного действия образуется разрыв длиной примерно в 30 нуклеотидов [2].
Стадии восстановления ДНК, в которой образовался циклобутановый димер. Участок с этой структурой, мешающей копированию ДНК, распознают и вырезают эндонуклеазы семейства ХР (ксеродермы), а застраивает образовавшуюся брешь в 29 нуклеотидов полимераза d (или e) при наличии фактора репликации (RFC) и белка-помощника (PCNA). Заделывают такую большую брешь ДНК-полимеразы d или e, но им нужен помощник PCNA. Более того, для работы первого фермента необходим еще один белок - репликативный фактор С (RFC). В присутствии АТФ он связывается с концом цепи ДНК, который подходит к бреши с 5ў-стороны, затем присоединяется PCNA и образуется нестабильный тройной комплекс. После гидролиза АТФ конформация репликативного фактора меняется так, чтобы удерживать PCNA будто раскрытыми пальцами. Сам же этот помощник за счет временного размыкания своей кольцевой формы насаживается на дуплекс ДНК. К образовавшемуся стабильному комплексу присоединяется ДНК-полимераза d, которая и застраивает большую брешь. Сходным образом работает комплекс с участием ДНК-полимеразы e. Как видно, в устранении объемных повреждений участников много, да и сам процесс довольно сложен.
Если в клетках гены активно транскрибируются (т.е. синтезируется матричная РНК), восстановление по типу вырезания нуклеотида протекает намного быстрее благодаря включению в процесс еще двух белков - CSA и CSB. У человека мутации в генах этих белков вызывают наследственное заболевание - синдром Кокэйна (Cockayne’s syndrom, CS), при котором замедляется рост, повышается фоточувствительность, возникают катаракта, кариес зубов и дерматозы. Если во время транскрипции осуществляющая ее РНК-полимераза II наталкивается на повреждение в ДНК, то связывается с белками CSA и CSB. Тогда быстрая доставка ферментов репарации к месту поломки гарантирована, они залатают прореху, после чего транскрипция может завершиться.
Ткацкие ошибки. Ошибки репликации и рекомбинация (обмен участками) между аллелями могут привести к появлению неспаренных нуклеотидов в цепи ДНК. Для устранения таких неполадок существует особый тип репарации - исправление ошибок спаривания (mismatch repair, MMR). Самый простой путь их коррекции - немедленно удалить неправильно встроенный нуклеотид с помощью все тех же ДНК-полимераз d или e (правда, с дополнительной - экзонуклеазной - активностью). Более сложный вариант осуществляют, совместно действуя, ферменты, которые способны вырезать поврежденный участок (ферменты эксцизионной репарации), и группа дополнительных белков. Они узнают одиночный неспаренный нуклеотид или петли длиной до четырех нуклеотидов. (Мутации генов этих белков у человека вызывают предрасположенность к раковым заболеваниям.) Как только дефект обнаружен, нуклеазы удаляют его вместе с 50-500 нуклеотидами с каждой стороны [3]. Огромную брешь застраивают те же полимеразы, зависимые от PCNA и потому способные синтезировать длинные фрагменты ДНК.
Регуляция. PCNA (о нем уже шла речь как о помощнике в репарации) и белок 53 (р53) - главные регуляторы в клетке, ответственные за ее судьбу. Первый белок, PCNA, участвует в репликации, второй - р53 - в транскрипции. А что делают эти белки, если во время копирования ДНК возникло повреждение, устранить которое должна ДНК-полимераза d? Белок 53 стимулирует синтез белка 21, а тот тормозит копирование ДНК и прохождение клеточного цикла. За время остановки репликации клетка успевает избавиться от дефекта. Как такое может быть, ведь репликация и репарация - это всегда синтез ДНК, в котором задействован один и тот же белок PCNA? Репликация останавливается из-за того, что р21 связывается с PCNA, значит, на репарацию его не будет хватать. Но нет: в области повреждения создается избыток PCNA, и оно успешно ликвидируется.
Некоторые ученые полагают, что при повышенном содержании р21 ресинтез ведет не полимераза d, а полимераза b, которая не чувствительна к действию р21. Мы же, исходя из наших результатов, предполагаем, что смены ферментов не происходит, а просто ДНК-полимераза d переключается с зависимого от PCNA заполнения брешей на независимый от этого белка синтез ДНК [1].
Когда структура ДНК восстановлена, ряд белков-регуляторов и ферментов, участвовавших в репарации, подвергаются гидролизу, который осуществляет 26S протеасома [4]. К таким белкам относится, например, белок XPC. Кроме того, сама протеасома или входящий в ее состав регуляторный комплекс 19S могут играть роль молекулярных шаперонов, способствуя приобретению репаративными белками нужной конформации. В этом случае протеасома стимулирует коррекцию ДНК.
Сквозные разрывы
Сквозные разрывы в молекуле ДНК - это результат ионизирующего облучения, окисления или механического повреждения. Такие же поломки возможны в тех случаях, когда во время репликации на пути ДНК-полимераз встречается разрыв в одной цепи. Но двухцепочечные разрывы могут быть и промежуточными продуктами нормальных биологических процессов (например, рекомбинации в развивающихся лимфоидных клетках). Если клетка не может заштопать сквозные разрывы, это приведет к дестабилизации генома, мутациям и возникновению раковых опухолей, а иногда к запуску апоптоза (запрограммированной гибели клетки). У эукариот существует два основных способа устранить двухцепочечные разрывы: гомологичная рекомбинация (рекомбинационная репарация) и соединение негомологичных концов.
Первый способ восстановления ДНК характерен для дрожжей (Saccharomyces cerevisiae), второй - для млекопитающих, в том числе для человека.
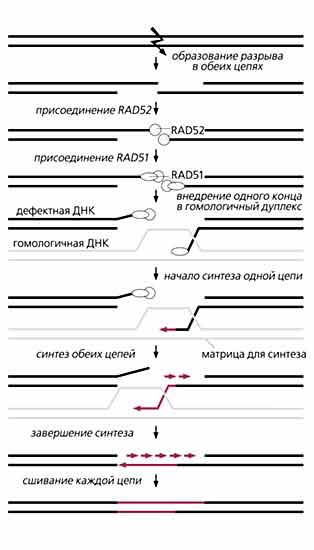
Объяснения в тексте.Гомологичный обмен. Белок RAD52, от которого зависит рекомбинационная репарация у дрожжей, связывается, как только что упоминалось, с выступающими концами разорванных цепей ДНК, чем защищает их от действия экзонуклеаз. Затем RAD52 стимулирует присоединение белка RAD51 к тем местам, где находился сам. После этого один конец или оба внедряются в гомологичный дуплекс ДНК и происходит рекомбинация. Выступавшие из “прорехи” части ДНК вырезаются, и образовавшаяся брешь застраивается сразу с противоположных сторон, т.е. синтез фрагментов идет навстречу друг другу. В клетках дрожжей S.cerevisiae его осуществляют полимеразы d и e, которые нуждаются в PCNA и репликативном факторе (RFC), чтобы удержаться на оборванном конце ДНК и вести синтез. Правда, в застраивании бреши участвует еще и ДНК-полимераза a, единственная из многочисленных полимераз, способная инициировать синтез ДНК в репликативной вилке - там, где начинают расплетаться нити. Учитывая участие этого фермента в репарации сквозных разрывов ДНК, А.Холмс и Дж.Хэйбер предложили модель, по которой внедрение одного из разорванных концов в гомологичный дуплекс ДНК создает необычную репликативную вилку, где и синтезируются цепи [6]. Репликация завершается, когда второй конец достигает разрыва. Оказавшийся лишним “повисший” концевой участок второй цепи удаляется нуклеазой.
С гомологичной рекомбинацией связан еще один способ латания мест разлома. Разорванные концы ДНК по кусочку отрезаются специфической экзонуклеазой в обоих направлениях от разрыва до тех пор, пока не откроются комплементарные последовательности. Затем следует отжиг (спаривание) комплементарных участков и подрезание “висящих” негомологичных хвостов ДНК. Этот путь репарации также зависит от белка RAD52.
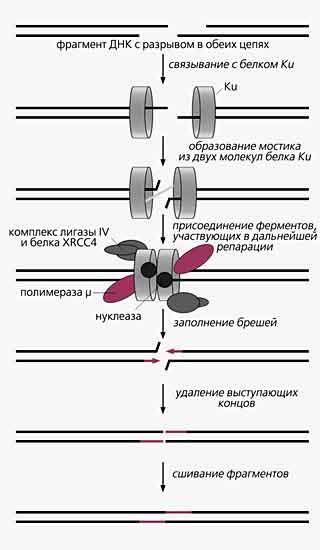
Соединение негомологичных концов. Починка этим способом ДНК, разорванной в обеих цепях, обеспечивается целым набором белков. Это белок Ku и комплекс, образованный ДНК-лигазой IV и продуктом гена XRCC4 [7]. Все они консервативны у эукариот, включая дрожжи и млекопитающих. Несмотря на то, что в ходе репарации порванные концы соединяются напрямую, без использования матрицы для синтеза, процесс должен быть максимально аккуратным и точным. Для выполнения этого требования служит белок Ku, который представляет собой гетеродимер из субъединиц Ku70 и Ku80 (молекулярная масса 70 и 80 кДа соответственно). Недавно была исследована кристаллическая структура человеческого гетеродимера, а также структура его комплекса с фрагментом ДНК в 55 нуклеотидов. Оказалось, что гетеродимер Ku кольцом обвивается вокруг дуплекса ДНК, но при этом не контактирует с основаниями ДНК, а образует несколько связей с сахарофосфатным остовом и подгоняется пространственно к виткам спирали ДНК так, чтобы они расположились в кольце белка строго определенным образом. Такая конфигурация комплекса Ku и ДНК, очевидно, необходима для поддержания той структуры выступающих оборванных концов, которая пригодна для следующих этапов репарации.
А их несколько. Сначала на оборванные концы ДНК “надевается” по гетеродимеру, затем они связываются друг с другом, образуя мостик. Именно к нему присоединяются и выполняют свои функции остальные участники: ДНК-полимераза заполняет бреши, образовавшиеся в результате соединения негомологичных концов ДНК; нуклеаза подрезает выступающие концы, если они оказались излишне длинными; лигаза IV сшивает восстановленные фрагменты в единое целое. У некоторых эукариот обнаружена ДНК-полимераза m, которая, видимо, и заполняет бреши в ДНК.
Когда сшиты цепи
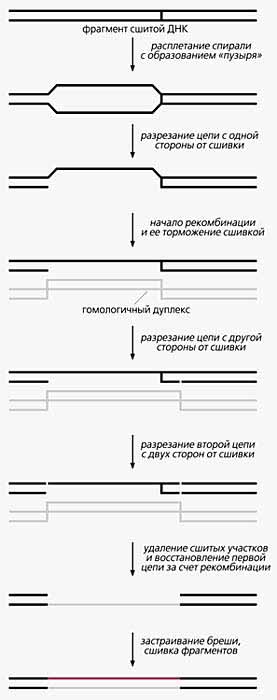
Некоторые химические агенты, например, лекарства против рака (цисплатин, митомицин, псорален), вызывают образование межцепочечных сшивок ДНК [8]. Такие неполадки эукариоты способны устранять, но механизмы репарации недостаточно ясны. Известно, что у почкующихся дрожжей S.cerevisiae она зависит от ферментных систем, обеспечивающих, например, удаление тиминовых димеров (о чем уже было сказано), и от рекомбинации. Эта же ферментная система из семи белков (ХРА-XPG) работает, как мы уже отмечали, и при исправлении человеческой ДНК со сшитыми цепочками. Только происходит все еще сложнее. Вспомним: один из белков, XPF, разрезает цепь ДНК с 5ў-стороны от повреждения на расстоянии 20(±5) нуклеотидов, другой, XPG, - с 3ў-стороны через 6(±3) нуклеотидов от димера. Однако в ДНК человека эти эндонуклеазы вырезают 22-28 нуклеотидов в одной цепи ДНК и лишь с одной стороны от сшивки, не затрагивая сам дефект. Почему же человеческая нуклеаза не разрезает цепь обычным образом, с обеих сторон от повреждения? Потому, полагают исследователи, что перед разрезанием двойная спираль должна быть расплетена (для этого служит фермент геликаза). Но расплести дуплекс она может не по соседству со сшивкой, а на отдалении от нее - на расстоянии 20(±5) нуклеотидов с 5ў-стороны. Расплетенный участок приобретает вид пузыря. Тогда-то одна эндонуклеаза, XPG, рвет цепь на положенном ей месте (т.е. ближе к дефекту), а другая, XPF, - приблизительно 27-ю фосфодиэфирную связь с 5ў-стороны от сшивки [9]. Но это всего лишь первый этап репарации ДНК с межцепочечными сшивками.
Что же дальше? По одной из разработанных гипотетических моделей, дальше следует рекомбинация гомологичного дуплекса ДНК. Именно большая брешь в 22-28 нуклеотидов инициирует этот процесс. Но на стадии переноса цепи он блокируется, после чего специфическая эндонуклеаза разрезает ту же нить с другой стороны сшивки. Вновь образовавшийся интермедиат узнают компоненты “молекулярных ножниц” (системы эксцизионной репарации), и осуществляется второй раунд разрезания, теперь уже двойного, во второй нити. В результате высвобождаются сшитые олигомеры, и рекомбинация завершается. Таким образом, одна цепь оказывается репарированной рекомбинацией. Брешь во второй цепи застраивается ДНК-полимеразой с использованием первой в качестве матрицы.
Интересно отметить, что недавно у высших эукариот обнаружен белок, содержащий домены геликазы и ДНК-полимеразы. Мутации его гена значительно повышают чувствительность организма к агентам, вызывающим сшивки между цепочками ДНК. А поскольку и геликазная, и полимеразная активности необходимы на определенных этапах восстановления сшитой ДНК за счет рекомбинации, был сделан вывод об участии этого белка в таком репарационном процессе. Впоследствии уникальный белок назвали ДНК-полимеразой q.
Гипотетическая схема устранения межцепочечных сшивок ДНК.
Объяснения в тексте.Если повреждение не удалено
Итак, повреждения ДНК у эукариот бывают разнообразными. И хотя существуют эффективные механизмы устранения таких дефектов, иногда клетка не способна удалить их все до единого по тем или иным причинам. В этом случае на поврежденных участках матрицы репликация ДНК останавливается. До недавнего времени было непонятно, каким же образом осуществляется копирование ДНК на поврежденных участках, не устраняемых известными способами. Это выяснилось совсем недавно.
В последние несколько лет открыта новая группа ДНК-полимераз - z, h, k, i, m, REV1, - которая как раз и предназначена для избавления от подобных повреждений [10]. Полимеразы z, h и m осуществляют ресинтез ДНК, если в ней оказались неустраненными циклобутановые димеры между соседними тиминами; два последние фермента, а также полимеразы k и REV1 - при сохранении окисленного гуанина и апуриновых сайтов, а полимераза REV1 успешно застраивает еще и места с О(6)-метилгуанином. Интересно, что z-полимераза “работает в паре” с полимеразой h или REV1, продлевая на несколько нуклеотидов пройденный ими поврежденный участок ДНК.
Перечисленные полимеразы во время репликации ДНК иногда встраивают правильные нуклеотиды против поврежденных участков матрицы, при этом структура ДНК не нарушается. В некоторых же случаях включаются ошибочные нуклеотиды, и тогда возникают мутации. В настоящее время новой группе ДНК-полимераз уделяется особое внимание, так как клеткам необходим энзиматический аппарат, способный синтезировать ДНК в ситуациях, когда репарация вырезанием неэффективна или невозможна, а риск накопления мутаций оправдан.
Первоначально механизм, предназначенный для устранения повреждений в ДНК, видимо, возник вместе с клеткой. По мере эволюции организмов он усложнялся, появлялись новые варианты. Насколько важен такой механизм, можно судить по тому, что в устранении поломок ДНК задействовано несколько десятков генов, и на производство ферментов репарации клетка тратит большую часть своих ресурсов. Надежность сохранения нуклеотидных последовательностей ДНК у высших эукариот очень велика: например, в геноме млекопитающего, насчитывающем около 3 млрд пар оснований, в клетках зародышевого пути за год подвергаются заменам лишь 10-20 оснований, а ведь повреждаются тысячи нуклеотидов. Особой надежности требует эмбриональный период: когда клетки активно делятся, необходимо как можно быстрее починить многочисленные поломки и сохранить генетическую информацию неповрежденной. Системы, благодаря которым восстанавливается природная структура ДНК, не только защищают геном от всевозможных повреждений, но и обслуживают генетические процессы в клетке. И если по какой-либо причине нарушен сам процесс репарации, это может обернуться серьезными неприятностями для клеток и всего организма. Так возникают повышенная предрасположенность к тем или иным заболеваниям, вплоть до онкологических, и тяжелые наследственные болезни. Знание механизмов, приводящих к различным заболеваниям, крайне необходимо практической медицине, чтобы облегчить поиск путей их лечения. И уж конечно, знание устраняет очередной пробел в постижении мира, будь он даже молекулярным. * * *
Работа поддержана Российским фондом фундаментальных исследований. Проекты 00-04-49183, 03-04-49127.
Литература
1. Sharova N.P., Abramova E.B., Dmitrieva S.B., et al. // FEBS Lett. 2000. V.486. P.14-18.
2. Sancar A. // Annu. Rev. Biochem. 1996. V.65. P.43-81.
3. Wood R.D., Shivji M.K.K. // Carcinogenesis. 1997. V.18. P.605-610.
4. Абрамова Е.Б., Шарова Н.П., Карпов В.Л. // Мол. биология. 2002. Т.36. С.761-776.
5. Sekelsky J.J., Burtis K.C., Hawley R.S. // Genetics. 1998. V.148. P.1587-1598.
6. Holmes A.M., and Haber J.E. // Cell. 1999. V.96. P.415-424.
7. Haber J.E. // Trends Genet. 2000. V.16. P.259-264.
8. Kohn K.W. // Cancer Res. 1996. V.56. P.5533-5546.
9. Besshno T., Mu D., and Sancar A. // Mol. Cell. Biol. 1997. V.17. P.6822-6830.
10. Burgers P.M.J., Koonin E.V., Bruford E., et al. // J. Biol. Chem. 2001. V.276. P.43487-43490.