
№12, 2001 г.
© И.А. ШевелевВОЛНОВЫЕ ПРОЦЕССЫ
В ЗРИТЕЛЬНОЙ КОРЕ МОЗГАИ.А.Шевелев
Игорь Александрович Шевелев, академик РАН,
директор Института высшей нервной деятельности и нейрофизиологии РАН.
В 1920-х годах немецкий электрофизиолог и психиатр Г.Бергер (1873—1941) впервые зарегистрировал биоэлектрическую активность (в виде колебаний потенциалов) мозга человека. С тех пор приложено много усилий, чтобы понять, имеют ли такие колебания какое-то функциональное значение или всего лишь сопутствуют нейронной активности и служат индикатором состояния мозга. Сегодня мы знаем, что работа этого важнейшего органа человека основана на передаче электрических сигналов, которые одновременно генерируются множеством нервных клеток. Электрическую активность всего мозга можно представить как шум огромной толпы, миллиарды членов которой разговаривают одновременно. Ни у кого не вызывает сомнений, что голос отдельного нейрона важен в этом общем шуме — ведь именно нервная клетка участвует в анализе и переработке информации. А вот относительно всей многоголосицы ясности нет до сих пор.
Суммарная электрическая активность мозга (шум огромной толпы) записывается с помощью электроэнцефалографа, который регистрирует колебания электрических потенциалов в нескольких частотных диапазонах, или ритмах. Все вместе они формируют электроэнцефалограмму (ЭЭГ), анализируя которую, нейрофизиологи пытаются понять, как работает мозг.
Здесь речь пойдет только об одной его функции — зрительной — и об одном виде электрической активности, отраженной на ЭЭГ, о так называемом альфа-ритме, т.е. колебаниях электрических потенциалов с частотой от 8 до 13 Гц. В этом диапазоне частота ритма индивидуальна, но есть он практически у каждого человека и особенно мощно проявляется в зрительной области (она находится в затылочной части обоих полушарий) коры большого мозга в состоянии спокойного бодрствования с закрытыми глазами.
Альфа-ритм, как и электрическая активность мозга в диапазоне других частот, многие годы был предметом фундаментальных исследований, но теперь перекочевал в прикладные и полуприкладные работы. Это связано, видимо, с тем, что большинство подходов к анализу его роли исчерпали себя и оказались непродуктивными. Несмотря на множество специальных руководств и ряд журналов, заполненных статьями по ЭЭГ, значение альфа-ритма для работы мозга до сих пор дискуссионно, если не сомнительно.
Неясны и механизмы генерации этого ритма. Так, все еще преобладает точка зрения, что альфа-ритм — это колебания потенциалов, синхронно развивающиеся во всей зрительной коре, т.е. стоячая волна. Между тем много лет назад американские исследователи — математик У.Питс и невролог У.Мак-Каллок — предложили гипотезу сканирования, связанную именно с этим ритмом [1]. По их мнению, альфа-ритм отображает сканирование зрительной коры, считывание с нее информации движущейся волной возбуждения. Напомним, что в первичной проекционной области зрительной коры, куда приходят сигналы из сетчатки глаза через подкорковый зрительный центр, соблюдается так называемая ретинотопия. Это означает, что видимый мир, спроецированный на сетчатку (ретину) оптической системой глаза, отображается в коре принципиально такой же картиной возбуждения. Недаром зрительную кору называют корковой экранной структурой.
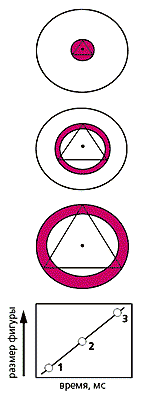
Распространение альфа-волны по зрительной коре. Согласно гипотезе Питса и Мак-Каллока, эта волна начинает свой ход (фаза 1) из центра корковой проекции поля зрения (ц.п.з.) к периферии. Однако экспериментальные результаты показывают обратное направление — от периферии к центру.Авторы гипотезы сканирования предположили, что через каждые 100 мс в центре этого коркового экрана возникает волна возбуждения, которая распространяется за такое же время до его границ. Эта движущаяся волна суммируется в каждом нейроне с возбуждением, пришедшим из сетчатки глаза, тем самым повышая уровень деполяризации нервных клеток, расположенных все дальше от центра зрительной коры. В результате с корковой экранной структуры последовательно считывается, сканируется, зрительная информация, которая передается затем в другие области коры, где производится дальнейший анализ информации и опознание образов. Авторы придавали весьма большое значение альфа-ритму и без излишней скромности назвали свою статью “Как мы познаем Вселенную. Восприятие слуховой и зрительной информации” *.* Позже, через 10 лет, Питс и Мак-Каллок стали соавторами знаменитой статьи Дж.Леттвина и Т.Матурано “Что глаз лягушки рассказывает мозгу лягушки”, в которой были впервые описаны нейроны-детекторы. К сожалению, это замечательное достижение не удостоилось Нобелевской премии вместе с Д.Хьюбелом и Т.Визелом (о них см.: Лауреаты Нобелевской премии 1981 года. По медицине — Р.Сперри, Д.Хьюбел, Т.Визел // Природа. 1982. №1. С.105—107).Мысль о сканирующей волне возникла не на пустом месте. Авторы заметили сходство структуры основных пучков нервных волокон в зрительной и слуховой коре со схемами устройств для автоматического управления зенитным огнем, разработанных в конце второй мировой войны. На это интересное совпадение обратил внимание и отец кибернетики Н.Винер, давший теоретической работе Питса и Мак-Каллока высокую оценку.Несомненно, что эта идея, окажись она верной, была бы очень важна как для понимания последовательности переработки зрительной информации, так и для оценки функционального значения альфа-ритма. Однако строгие доказательства гипотезы отсутствовали. Экспериментально сам сканирующий процесс не был обнаружен, а косвенные данные оказались противоречивыми — одни из них подтверждали следствия, вытекающие из гипотезы, другие — нет.
Чтобы подтвердить или опровергнуть гипотезу Питса и Мак-Каллока, в середине 80-х годов мы начали специальные опыты.
Четыре косвенных доказательства
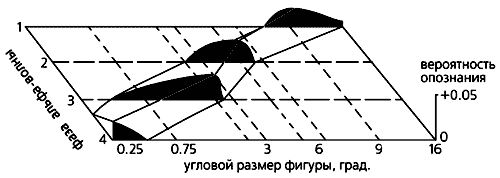
Опознание формы геометрических фигур. В то время проверить гипотезу об альфа-сканировании можно было лишь косвенно, так как существовало много методических ограничений. Мы провели 285 управляемых компьютером экспериментов, в которых перед испытуемыми (их было 29 человек) ставилась задача опознать форму геометрических фигур разного углового размера. Мы исходили из простого следствия гипотезы: если альфа-ритм распространяется из центра зрительной коры, то мелкие изображения, контур которых там и проецируется, будут опознаваться лучше при их предъявлении испытуемому на ранних фазах альфа-волны, а более крупные, “локализованные” на периферии, — на относительно поздних *.
* Точные фазы волны в то время было невозможно установить, поскольку сигналы от изображений разного размера приходят в зрительную кору с разной задержкой. Но это не имело принципиального значения для наших опытов, поэтому здесь говорится только о последовательности фаз альфа-волны — относительно ранних или относительно поздних.В экспериментах компьютер в случайном порядке, на короткое время, предъявлял на экране геометрические фигуры разной формы и углового размера (от 0.7 до 16°) во время одной из четырех фаз альфа-волны или же — в контрольной серии — вне всякой связи с какой-либо из них.Как мы и предполагали, вероятность опознания фигуры увеличивалась, если ее запуск производился в соответствующую ее размеру фазу [2, 3]. Но что было неожиданным, так это противоположная предсказанной связь между размером фигуры и фазой волны: если изображения на экране возникали в ранние фазы, лучше опознавались более крупные фигуры, а в поздние — наоборот, мелкие.
Таким образом, мы косвенно подтвердили следствие гипотезы Питса и Мак-Каллока о распространении волны возбуждения по зрительной коре. Однако из наших опытов следовало, что альфа-волна движется не от центра к периферии корковой проекции поля зрения, как предполагали авторы гипотезы, а наоборот — от периферии к центру. Мы полагаем, что каждые 80—120 мс (период альфа-ритма у разных испытуемых) в корковой проекции ближней периферии поля зрения (9—12° от его центра) генерируется волна, которая равномерно перемещается к центру и достигает его за три четверти альфа-периода, т.е. примерно за 75 мс.
Распознавание направления движения стимула. Естественно было предположить, что с фазами альфа-волны связано распознавание не только формы изображения, но и направления его движения. Чтобы выяснить, так ли это, вызванная быстро движущимся по экрану световым пятном волна коркового возбуждения пускалась либо в одном направлении с распространяющейся альфа-волной, либо — навстречу ей. Испытуемые должны были ответить, к центру экрана или к периферии движется световое пятно, создаваемое компьютером. В этой серии экспериментов соблюдались те же условия: в опыте начало движения зрительного стимула совпадало с одной из фаз альфа-ритма, в контроле синхронизация полностью отсутствовала.
Известно, что человек лучше опознает центростремительное направление. (Это связано, по-видимому, с большей биологической значимостью для человека сигналов, которые появляются на периферии поля зрения в связи с их новизной и возможной опасностью.) В контроле так и было. Но в опыте это предпочтение сменялось на противоположное: испытуемые лучше распознавали центробежное направление — навстречу альфа-волне [4, 5]. Такое предпочтение в опознании вполне объяснимо. Известно, что восприятие движения основано на активации нейронов зрительной коры, обладающих высокой чувствительностью к его направлению. А поскольку активация зависит от пространственно-временного градиента входного сигнала, который бывает наибольшим при встречном движении двух волн (центростремительной альфа-волны и центробежной волны, вызванной светом), то естественно, что именно в этом случае и улучшаются показатели распознавания. Так подтвердилось еще одно следствие гипотезы Питса и Мак-Каллока.
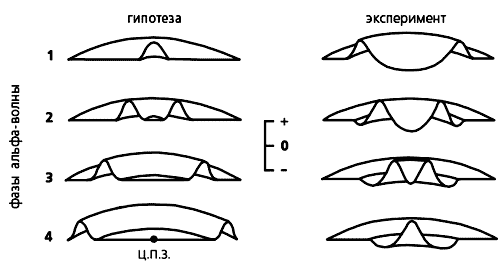
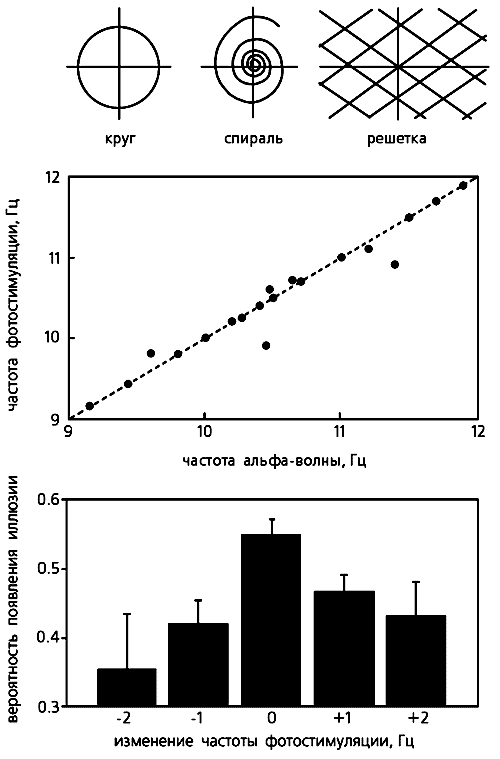
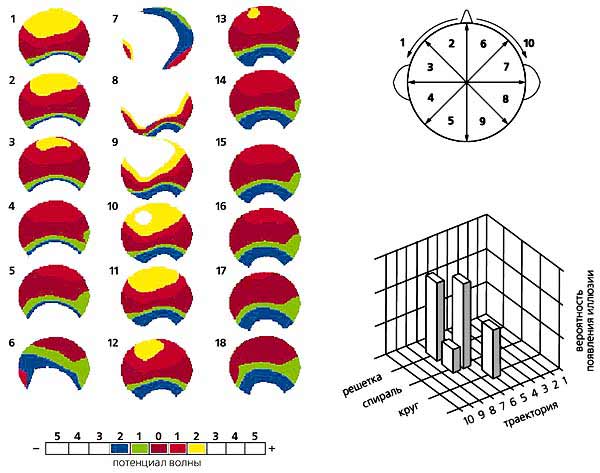
Зависимость вероятности (отложена по вертикали) опознания геометрических фигур разного размера от момента их предъявления, т.е. от фазы альфа-волны. При хаотическом предъявлении в контрольных опытах эта вероятность одинакова и принята за ноль (координатная плоскость). Видно, что распознавание более крупных фигур улучшается по сравнению с контролем, если они предъявляются на ранних фазах альфа-ритма, а более мелких — на поздних фазах.Возникновение зрительных иллюзий. Из гипотезы сканирования вытекает еще одно следствие: ритмическая фотостимуляция с частотой альфа-ритма может создать в зрительной коре квазистробоскопический эффект. Мы решили проверить, способны ли ритмические вспышки света, создающие в зрительной коре диффузную волну возбуждения, “заморозить”, остановить альфа-волну в некий момент движения и тем вызвать ее зрительное восприятие как неподвижного изображения. Следует сказать, что еще в 50-х годах английский нейрофизиолог У.Уолтер описал появление некоторых иллюзорных зрительных образов при вспышках света.В наших опытах человек сидел с закрытыми глазами перед лампой стробоскопа, частоту вспышек которого компьютер менял в случайном порядке. При возникновении зрительной иллюзии испытуемый нажимал на ключ и компьютер фиксировал частоту, на которой образ появлялся. Судя по словесным описаниям и зарисовкам восприятий, испытуемые видели светящиеся кольца, круги, спирали или решетки с косо пересекающимися линиями [6, 7]. Примечательно, что в многократных повторах у каждого испытуемого преобладал один и тот же тип иллюзии. Раз возникнув, она могла двигаться: спираль или диск медленно вращались, образ пульсировал, удаляясь или смещаясь в сторону, менял цвет.
Индивидуальная, доминирующая в спектре мощности ЭЭГ, частота альфа-ритма и оптимальная частота стимуляции (ее диапазон у разных людей колебался от 6.5 до 13.8 Гц) для получения четкого иллюзорного эффекта оказались тесно связанными. Характерно, что вероятность восприятия круга и спирали значимо снижалась даже при небольшом изменении частоты стимуляции, а образ решетки возникал на частотах более низких или высоких по сравнению с теми, которые вызывали иллюзии круга и спирали.
Итак, в этой серии опытов подтвердилось третье следствие исходной гипотезы. Наши испытуемые “увидели”, т.е. иллюзорно восприняли как реальный зрительный образ собственную волну альфа-ритма, как бы остановленную в тот или иной момент ее распространения по зрительной коре.
На чем основан этот эффект? Можно думать, что возбуждение, создаваемое вспышками, изоритмичными с альфа-волной, доводит до порога восприятия обычно невидимое (подпороговое) ее распространение. Наши результаты свидетельствуют в пользу кольцевой формы сканирующей волны, так как именно круг или кольцо были наиболее характерными иллюзиями у испытуемых. Что касается спирали, то она чаще возникала при небольшом рассогласовании частот стимуляции и альфа-ритма, в то время как решетка — при более значительном их отличии.
Движущиеся волны и зрительные иллюзии. Мы решили проверить также, связаны ли описанные иллюзии с движущимися волнами ЭЭГ, так как казалось логичным, что и эти волны могут отражать работу сканирующего механизма. Дело в том, что движение волны возбуждения в зрительной коре может вызвать последовательное смещение максимума потенциала по поверхности коры большого мозга. В электрофизиологии это смещение и принято называть движущимися волнами. На динамических картах альфа-потенциалов мозга удается выделить несколько фокусов, или полюсов, каждый из которых перемещается по своей траектории. Если проанализировать такие карты, можно выяснить, совпадает ли с каким-либо направлением движения та или иная иллюзия, вызванная у испытуемых ритмической фотостимуляцией.
Возникновение зрительных иллюзий. При фотостимуляции с частотой альфа-ритма обычно появляются иллюзии в виде круга, спирали или решетки (вверху), причем четко проявляется связь частоты фотостимуляции для их вызова с доминирующей у каждого испытуемого частотой альфа-ритма (в середине). Примечательно, что рассогласование этих частот на 1—2 Гц приводит к снижению вероятности возникновения любой иллюзии, особенно — круга (внизу).Карты альфа-потенциалов (с шагом 4 мс) мы строили на основе ЭЭГ испытуемых, которые находились в условиях наибольшего проявления альфа-ритма, т.е. спокойно сидели закрыв глаза. Движущиеся волны обнаруживали, сравнивая ЭЭГ, полученные до и во время фотостимуляции, и ранжировали их по 10 траекториям. Каковы же были результаты?Во-первых, на серии динамических карт, отражающих волну, которая двигалась от затылка ко лбу, мы увидели, что отрицательный полюс потенциала, находящийся в затылочной области коры, распространяется в том же направлении [8]. Во-вторых, удалось выявить, что конкретные иллюзии возникали на фоне определенной траектории волны. Например, круг и спираль появлялись в том случае, если волна двигалась от затылка ко лбу, а решетка — при траектории, направленной от левой затылочной области коры к правой лобной. Следовательно, связь между видом иллюзии и направлением движения альфа-волны существует.
Прямое доказательство
Теперь мы уже знали, что от фазы альфа-волны зависит распознавание формы геометрических фигур и направления движения зрительного стимула, что эту волну можно “остановить”, вызвав ее иллюзорный зрительный образ; увидели, что она распространяется по нескольким траекториям, с которыми связан определенный вид иллюзии. Но все это — косвенные доказательства. Как же получить прямые подтверждения теоретическим построениям Питса и Мак-Каллока?
Исходя из зависимости вида иллюзии от траектории смещения максимума потенциала, мы предположили, что эта траектория может отражать и движение источника альфа-ритма. Обнаружив это, мы тем самым непосредственно доказали бы гипотезу сканирования.
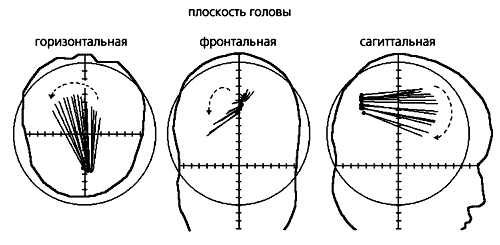
Опознание центробежного и центростремительного движений. При совпадении времени запуска движущегося светового пятна с какой-либо фазой альфа-волны центробежное направление опознается лучше по сравнению с контролем (нулевая плоскость), а центростремительное — хуже. Предпочтение выражено сильнее, когда пятно начинает движение из левого полуполя зрения. Звездочками на схеме отмечены достоверные (р<0.05) отклонения в опознании.Регистрация электрической активности мозга от набора электродов, расположенных в разных участках головы человека, позволяет судить о распределении потенциалов на двумерной поверхности. Известно, однако, что в энцефалограмму существенный, а иногда и определяющий вклад вносят генераторы потенциалов из глубинных структур мозга. Обнаружить и установить местоположение таких генераторов позволяют методы трехмерного анализа ЭЭГ, основанные на решении обратной задачи. Для этого разработаны специальные компьютерные программы.Считается, что за распределение альфа-потенциалов по поверхности головы отвечает эквивалентный токовый диполь альфа-ритма [9]. Наша задача состояла в том, чтобы выявить последовательный сдвиг диполя по зрительной коре во время развития альфа-волны, которая регистрируется от поверхности головы.
Для этого анализировали ЭЭГ испытуемых (в опытах с ритмической фотостимуляцией, вызывающей зрительные иллюзии), решая обратную задачу для трехслойной сферической модели головы. В результате в затылочных долях мозга удалось выявить и последовательный сдвиг диполя, и поворот его вектора [10]. При движении альфа-волны от затылочной области к лобной источник альфа-ритма смещался в медиальном направлении и вниз, а его дипольный момент поворачивался против часовой стрелки в горизонтальной плоскости и по часовой стрелке — в сагиттальной. Эти результаты указывают на быстрое смещение источника альфа-ритма в области мозга, которая совпадает с первичной зрительной корой, и подтверждают гипотезу о ее сканировании распространяющейся волной.
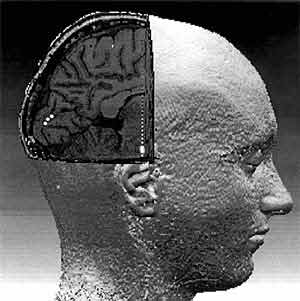
Движущаяся волна альфа-ритма и ее траектории. На картах поля альфа-потенциалов мозга (три левых колонки) одного из испытуемых видно, что она распространяется от затылочной области мозга к лобной. Возможны несколько траекторий (на правой части рисунка их показано 10). На фоне определенной траектории и возникает конкретная зрительная иллюзия (справа внизу).Таким образом, последовательный сдвиг диполя во время альфа-ритма прямо отражает движение волны возбуждения по зрительной коре. Поворот же вектора диполя при этом становится неизбежным, ведь она движется не по ровной поверхности, а по сложно упакованной шпорной извилине в первичной проекционной области зрительной коры.Поскольку локализация источника альфа-ритма проводилась на “обобщенной” трехмерной модели головы, мы сочли необходимым подтвердить его местоположение для каждого испытуемого с помощью ЯМР-томографии. Полученные трехмерные карты структур мозга (с пространственным разрешением около 1 мм) каждого конкретного человека, участвовавшего в опыте, совмещали по определенной программе с картами, которые отражали локализацию диполей альфа-ритма. Благодаря такому наложению двух видов трехмерных карт впервые удалось прямо выявить, что дипольный источник альфа-волны во время ее движения последовательно смещается по шпорной извилине. Это непосредственно, а не косвенно, свидетельствует в пользу гипотезы сканирования.
Положение токового диполя альфа-ритма и его векторов в динамике (шаг 2 мс). Токовый диполь (черные точки) — источник альфа-ритма — локализован в затылочной области, в какой бы плоскости профиля головы он ни был выявлен, и его позиция меняется по мере движения волны. Происходит и поворот вектора диполя (линии, исходящие из точек).Механизмы генерации и распространения альфа-волныТеперь, имея доказательства гипотезы Питса и Мак-Каллока, попробуем разобраться в механизмах, которые обусловливают возникновение и движение альфа-волны.
В принципе полученные в наших экспериментах эффекты можно было бы попытаться объяснить и без привлечения идеи о движении альфа-волны по зрительной коре. Однако если предположить, что одновременно вся эта кора просто “дышит” с частотой альфа-ритма, то нельзя было бы связать зависимость опознания геометрических фигур разного размера с фазой альфа-волны. Необъяснимым осталось бы и появление оформленных зрительных иллюзий при фотовспышках с альфа-частотой.
Часто критики гипотезы сканирования справедливо напоминают, что альфа-ритм наиболее выражен в темноте и при закрытых глазах, а воздействие света подавляет его, и он сменяется асинхронными колебаниями на более высоких частотах. Но противоречие этого известного факта с гипотезой Питса и Мак-Каллока только кажущееся, и авторы, кстати, сами упоминали о нем. Можно думать, что активация альфа-ритма при закрытых глазах — это эффект “холостого хода” сканирующего механизма. Тогда снижение на свету мощности альфа-ритма свидетельствует не об его истинном уменьшении, а об интерференционной маскировке. В самом деле, пришедшие из сетчатки глаза сигналы о многообразном внешнем мире по-разному активируют корковые нейроны, которыми эти сигналы (точнее, зрительная информация) перерабатываются. В результате создается множество волн возбуждения не синхронных с альфа-ритмом, потому и возникает впечатление, что он затухает при активном зрительном восприятии.
Питс и Мак-Каллок считали, что благодаря сканированию зрительной коры альфа-волной человек распознает объекты внешнего мира независимо от изменения их размера и поворота в пространстве. (Все мы узнаем стул, большой он или маленький, стоит на ногах или лежит на боку.) Кроме того, авторы гипотезы утверждали, что последовательное сканирование выгодно для представления на выходе зрительной коры информации не только в пространственном коде, но и во временном. К тому же сканирование, считали они, может обеспечить компактность “кабеля”, который связывает первичную зрительную кору с другими корковыми полями мозга. И этот “кабель” не требует того громадного числа нервных волокон, которое было бы необходимо для восприятия зрительной информации без сканирующего процесса. Благодаря ему, утверждали Питс и Мак-Каллок, такой “кабель” может быть упакован в отведенное ему ограниченное место в белом веществе затылочных долей мозга. Трудно добавить что-либо новое и существенное к этим весьма развитым умозрительным построениям, высказанным 50 лет назад. И хотя их все еще трудно обсуждать в деталях, аналогии можно найти. Отечественный физиолог М.Н.Ливанов (1907—1986) говорил о пользе сонастройки ритмов активности разных структур мозга для обеспечения функциональной связи между ними. За счет такой связи и повышается эффективность двигательной реакции на сенсорный стимул у животных и человека.
Точная локализация токовых диполей в мозге человека. На трехмерной карте мозга, полученной по данным ЯМР-томографии, расчетным методом виртуальной хирургии удалена часть затылочного полюса правого полушария. На внутренней, “обнажившейся” поверхности левого полушария показаны три последовательных положения дипольного источника альфа-ритма (белые точки). Находятся они на шпорной борозде — первичной проекционной области зрительной коры (другие дипольные источники располагаются глубже и на срезе не видны). Из-за сложности рельефа этой борозды поворот вектора диполя становится неизбежным.Известно, что волны возбуждения могут генерироваться как внутри коры мозга, так и в подкорке, т.е. в структурах, внешних по отношению к ней. В самой коре волны, движущиеся со скоростью нескольких метров в секунду (такова скорость и сканирующей волны), до сих пор прямо не визуализированы. Правда, благодаря применению все более совершенных технических устройств в нейрофизиологических исследованиях уже удалось увидеть более медленные, чем сканирующая альфа-волна, волны (со скоростями до нескольких десятков мм/c) в зрительной коре [11] и обонятельной луковице [12]. Мы надеемся, что удастся увидеть, а не иллюзорно воспринять, и движущуюся альфа-волну. Мы уверены, что ее визуализация позволит лучше понять последовательность операций по переработке зрительной информации в коре большого мозга.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проекты 93-04-21450, 97-04-48415), Российского гуманитарного научного фонда (проект 98-06-08027а), а также программы “Интеграция” (проект 429).Литература 1. Pitts W., McCulloch W.S. // Bull. Math. Biophys. 1947. V.9. P.127—147.
2. Шевелев И.А. и др. // Физиология человека. 1985. Т.11. №5. С.707—711.
3. Shevelev I.A. et al. // Int. J. Psychophysiol. 1991. V.11. P.195—201.
4. Шевелев И.А. и др. // Сенсор. системы. 1991. Т.5. №3. С.54—59.
5. Shevelev I.A. et al. // Febs Lett. 1996. V.392. P.169—174.
6. Шевелев И.А. и др. // Журн. высш. нерв. деятельности. 1996. Т.46. №1. С.34—39.
7. Каменкович В.М. и др. // Журн. высш. нерв. деятельности. 1998. Т.48. №3. С.449—457.
8. Shevelev I.A. et al. // Int. J. Psychophysiol. 2000. V.39. P.9—20.
9. Верхлютов В.М. и др. // Журн. высш. нерв. деятельности. 1999. Т.49. №1. С.3—11.
10. Shevelev I.A. // Brain Topogr. 1993. V.5. №2. P.77—85.
11. Shevelev I.A., Tsicalov E.N. // Neuroscience. 1997. V.76. №2. P.531—540.
12. Delaney K.R. et al. // Proc. Nat. Acad. Sci. USA. 1994. V.91. P.669—673.


Декабрь 2001