|
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ
НАУК
том 73, № 9, с. 797-806 (2003) |
|
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ
НАУК
том 73, № 9, с. 797-806 (2003) |
К 50-летию открытия структуры ДНК
© А.В. Зеленин
ГЕНОМ РАСТЕНИЙ
А. В. Зеленин
Зеленин Александр Владимирович - д.б.н.,
заведующий лабораторией Института молекулярной биологии им. В.А. Энгельгардта РАН.Впечатляющие достижения программы "Геном человека", а также успехи работ по расшифровке так называемых сверхмалых (вирусы), малых (бактерии, дрожжи) и средних (круглый червь, дрозофила) геномов сделали возможным переход к широкомасштабному изучению крупных и сверхкрупных геномов растений [1]. Насущная необходимость детального изучения геномов наиболее важных в хозяйственном отношении растений была подчеркнута на совещании по геномике растений, состоявшемся в 1997 г. в США [2, 3]. За прошедшие с того времени годы достигнуты несомненные успехи в этой области. В 2000 г. появилась публикация о полном секвенировании (установление линейной последовательности нуклеотидов всей ядерной ДНК) генома горчицы малой - арабидопсиса, в 2001 г. - о предварительном (черновом) секвенировании генома риса. Неоднократно сообщалось о работах по секвенированию крупных и сверхкрупных геномов растений (кукуруза, рожь, пшеница), однако эти сообщения не содержали конкретной информации и носили, скорее, характер деклараций о намерении.
Предполагается, что расшифровка геномов растений откроет перед наукой и практикой широкие перспективы. Прежде всего выявление новых генов и цепочки их генетической регуляции позволит существенно повысить продуктивность растений за счет использования биотехнологических подходов. С обнаружением, выделением, размножением (клонированием) и секвенированием генов, отвечающих за такие важнейшие функции растительного организма, как размножение и продуктивность, процессы изменчивости, устойчивости к воздействию неблагоприятных факторов среды, а также гомологичное спаривание хромосом, связывают появление новых возможностей для усовершенствования селекционного процесса. Наконец, выделенные и клонированные гены можно использовать для получения трансгенных растений с принципиально новыми свойствами и анализа механизмов регуляции активности генов.
Важность изучения геномов растений подчеркивает и то обстоятельство, что до настоящего времени число локализованных, клонированных и секвенированных генов растений невелико и колеблется, по различным оценкам, между 800 и 1200. Это в 10-15 раз меньше, чем, например, у человека.
Несомненным лидером в широкомасштабном изучении геномов растений остаются США, хотя интенсивные исследования генома риса проводятся в Японии, а в последние годы и в Китае. В расшифровке генома арабидопсиса, кроме лабораторий США, приняли активное участие исследовательские группы Европы. Явное лидерство США вызывает серьезное беспокойство европейских ученых, которое они ясно выразили на совещании под многозначительным названием "Перспективы геномики в постгеномную эру", состоявшемся в конце 2000 г. во Франции. Опережение американской науки в изучении геномов сельскохозяйственных растений и создании трансгенных растительных форм, по мнению европейских ученых, грозит тем, что в не слишком отдаленном будущем (от двух до пяти десятилетий), когда рост численности населения поставит человечество перед лицом всеобщего продовольственного кризиса, европейская экономика и наука попадут в зависимость от американских технологий. В связи с этим объявлено о создании франко-германской научной программы по исследованию геномов растений ("Plantgene") и вложении в нее значительных средств.
Очевидно, что проблемы геномики растений должны привлечь пристальное внимание российских ученых и организаторов науки, а также руководящих инстанций, поскольку речь идет не только о научном престиже, но и о национальной безопасности страны. Через одно-два десятилетия продовольствие станет важнейшим стратегическим ресурсом.
ТРУДНОСТИ В ИЗУЧЕНИИ ГЕНОМОВ РАСТЕНИЙ
Изучение геномов растений - задача значительно более сложная, чем исследование генома человека и других животных [1]. Это связано со следующими обстоятельствами:
• огромными размерами геномов, достигающими для отдельных видов растений десятков и даже сотен миллиардов пар нуклеотидов (п.н.): геномы основных хозяйственно важных растений (кроме риса, льна и хлопка) по размерам либо близки к геному человека, либо превышают его во много раз (таблица);ХРОМОСОМНЫЕ ИССЛЕДОВАНИЯ ГЕНОМОВ• резкими колебаниями числа хромосом у различных растений - от двух у некоторых видов до нескольких сотен у других, причем не удается выявить строгой корреляции между размером генома и числом хромосом;
• изобилием полиплоидных (содержащих более двух геномов на клетку) форм с близкими, но не идентичными геномами (аллополиплоидия);
• чрезвычайной обогащенностью геномов растений (до 99%) "незначащей" (некодирующей, то есть не содержащей генов) ДНК, что резко затрудняет стыковку (расположение в правильном порядке) отсеквенированных фрагментов в общий крупноразмерный участок ДНК (контиг);
• неполным (по сравнению с геномами дрозо-филы, человека и мыши) морфологическим, генетическим и физическим картированием хромосом;
• практической невозможностью выделять в чистом виде индивидуальные хромосомы с помощью методов, обычно применяемых с этой целью для хромосом человека и животных (сортировка в потоке и использование гибридов клеток);
• трудностью хромосомного картирования (определение расположения на хромосоме) отдельных генов с помощью гибридизации in situ, обусловленной как высоким содержанием в геномах растений "незначащей" ДНК, так и особенностями структурной организации хромосом растений;
• эволюционной отдаленностью растений от животных, что серьезно осложняет использование для изучения геномов растений сведений, полученных при секвенировании генома человека и других животных;
• длительным процессом размножения большинства растений, что существенно замедляет их генетический анализ.
Хромосомные (цитогенетические) исследования геномов вообще и растений в частности имеют длинную историю. Термин "геном" был предложен для обозначения гаплоидного (единичного) набора хромосом с содержащимися в них генами в первой четверти XX в., то есть задолго до установления роли ДНК как носителя генетической информации [4].
Описание генома нового, ранее генетически не изученного многоклеточного организма обычно начинают с исследования и описания полного набора его хромосом (кариотипа). Это, разумеется, относится и к растениям, огромное множество которых еще даже не начали изучать.
Уже на заре хромосомных исследований проводили сравнение геномов родственных видов растений на основе анализа мейотической конъюгации (объединения гомологичных хромосом) у межвидовых гибридов. За прошедшие 100 лет возможности хромосомного анализа резко расширились. Сейчас для характеристики геномов растений используют более совершенные технологии: различные варианты так называемого дифференциального окрашивания, позволяющего по морфологическим признакам идентифицировать индивидуальные хромосомы; гибридизацию in situ, дающую возможность локализовать конкретные гены на хромосомах; биохимические исследования клеточных белков (электрофорез и иммунохимия) и, наконец, комплекс методов, основанных на анализе хромосомной ДНК вплоть до ее секвенирования.
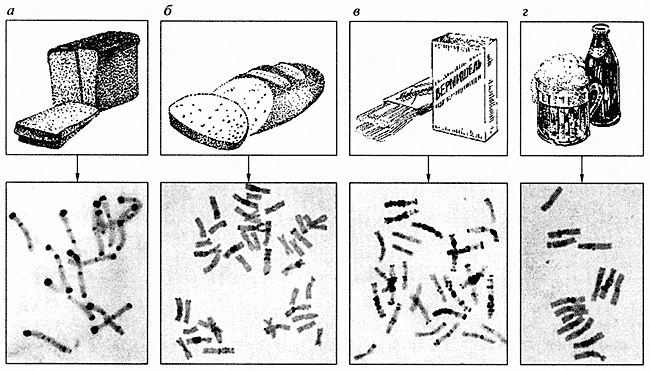
Рис. 1. Кариотипы хлебных злаков а - рожь (14 хромосом), б - твердая пшеница (28 хромосом), в - мягкая пшеница (42 хромосомы), г - ячмень (14 хромосом)В течение многих лет изучаются кариотипы хлебных злаков, прежде всего пшеницы и ржи. Интересно, что у разных видов этих растений число хромосом различно, но всегда кратно семи. Отдельные виды хлебных злаков можно надежно распознать по их кариотипу. Например, геном ржи состоит из семи пар крупных хромосом, имеющих на своих концах интенсивно окрашенные гетерохроматические блоки, часто называемые сегментами, или бэндами (рис. 1, а). Геномы пшеницы насчитывают уже 14 и 21 пару хромосом (рис. 1, б, в), причем распределение в них гетерохроматических блоков не такое, как в хромосомах ржи. Различаются между собой и отдельные геномы пшеницы, получившие обозначение А, В и D. Возрастание числа хромосом с 14 до 21 приводит к резкому изменению свойств пшеницы, что нашло отражение в их названиях: твердая, или макаронная, пшеница и мягкая, или хлебная, пшеница. За приобретение мягкой пшеницей высоких хлебопекарных свойств ответственен геном D, содержащий гены белков клейковины, которая придает тесту так называемую всхожесть. Именно этому геному уделяется особое внимание при селекционном улучшении хлебных пшениц. Другой 14-хромосомный злак - ячмень (рис. 1, г) обычно не используют для приготовления хлеба, однако он служит основным сырьем для изготовления таких распространенных продуктов, как пиво и виски.Интенсивно изучаются хромосомы некоторых дикорастущих растений, используемых для улучшения качества важнейших сельскохозяйственных видов, например диких сородичей пшеницы - эгилопсов [5]. Новые растительные формы создаются путем скрещивания (рис. 2) и отбора. В последние годы значительное усовершенствование методики исследований позволило приступить к изучению геномов растений, особенности кариотипов которых (главным образом мелкие размеры хромосом) делали их ранее недоступными для хромосомного анализа. Так, лишь недавно были впервые идентифицированы все хромосомы хлопка, ромашки и льна [6].

Рис. 2. Кариотипы пшеницы и гибрида пшеницы с эгилопсом
а - гексаплоидная мягкая пшеница (Triticum astivum), состоящая из А, В и О геномов; б - тетраплоидная пшеница (Triticum timopheevi), состоящая из А и G геномов. содержит гены устойчивости к большинству болезней пшеницы; в - гибриды Triticum astivum х Triticum timopheevi, устойчивые к мучнистой росе и ржавчине, отчетливо видно замещение части хромосомПЕРВИЧНАЯ СТРУКТУРА ДНКПо мере развития молекулярной генетики расширилось само понятие генома. Сейчас этот термин трактуется как в классическом хромосомном, так и в осовремененном молекулярном смысле: весь генетический материал отдельного вируса, клетки и организма [4]. Естественно, что вслед за изучением полной первичной структуры геномов (так часто называют полную линейную последовательность оснований нуклеиновых кислот) ряда микроорганизмов и человека на очередь встал вопрос о секвенировании геномов растений.
Из множества растительных организмов для исследования были выбраны два - арабидопсис, представляющий класс двудольных (размер генома 125 млн. п.н.), и рис из класса однодольных (420-470 млн. п.н.). Эти геномы невелики по сравнению с геномами других растений и содержат сравнительно немного повторяющихся участков ДНК. Такие особенности давали надежду на то, что выбранные геномы окажутся доступными для относительно быстрого определения их первичной структуры.
Рис. 3. Арабидопсис - горчица малая - мелкое растение из семейства крестоцветных (Brassicaceae). На пространстве, равном по площади одной странице нашего журнала, можно вырастить до тысячи индивидуальных организмов арабидопсисаОснованием для выбора арабидопсиса послужили не только небольшие размеры его генома, но и мелкие размеры организма, что позволяет легко выращивать его в лабораторных условиях (рис. 3). Принимали во внимание его короткий репродуктивный цикл, благодаря чему можно быстро проводить опыты по скрещиванию и отбору, детально изученную генетику, легкость осуществления манипуляций со сменой условий произрастания (изменение солевого состава почвы, добавление разных питательных веществ и т.д.) и с испытанием действия на растения различных мутагенных факторов и патогенов (вирусы, бактерии, грибы). Арабидопсис не имеет хозяйственной ценности, поэтому его геном, наряду с геномом мыши, получил название справочного, или, что менее точно, модельного.** Появление в отечественной литературе термина "модельный геном" - результат неточного перевода английского словосочетания model genome. Слово "model" означает не только прилагательное "модельный", но и существительное "образец", "эталон", "модель". Правильнее было бы говорить о геноме-образце, или справочном геноме.Интенсивная работа по секвенированию генома арабидопсиса была начата в 1996 г. международным консорциумом, в который вошли научные учреждения и исследовательские группы из США, Японии, Бельгии, Италии, Великобритании и Германии. В декабре 2000 г. стала доступной обширная информация, подводившая итоги определения первичной структуры генома арабидопсиса [7]. Для секвенирования использовали классическую, или иерархическую, технологию: сначала изучали отдельные небольшие участки генома, из которых составляли более крупные участки (контиги), а на финальном этапе - структуру индивидуальных хромосом. Ядерная ДНК генома арабидопсиса распределена между пятью хромосомами. В 1999 г. были опубликованы результаты секвенирования двух хромосом, а появление в печати сведений о первичной структуре остальных трех завершило секвенирование всего генома.Из 125 млн. пар нуклеотидов определена первичная структура 119 млн., что составляет 92% всего генома. Лишь 8% генома арабидопсиса, содержащих крупные блоки повторяющихся участков ДНК, оказались недоступными для изучения. По полноте и тщательности секвенирования геномов эукариот арабидопсис остается пока в первой тройке чемпионов наряду с одноклеточным дрожжевым организмом Saccharomyces cerevisiae и многоклеточным организмом животного Саеnorhabditis elegance (см. табл.).
В геноме арабидопсиса обнаружено около 15 тыс. индивидуальных генов, кодирующих белки. Приблизительно 12 тыс. из них содержатся в виде двух копий на гаплоидный (единичный) геном, так что общее число генов составляет 27 тыс. Число генов у арабидопсиса не сильно отличается от числа генов у таких организмов, как человек и мышь, однако размеры его генома в 25-30 раз меньше. С этим обстоятельством связаны важные особенности в структуре отдельных генов арабидопсиса и общей структуры его генома.
Гены арабидопсиса компактны, содержат лишь несколько экзонов (участков, кодирующих белки), разделенных короткими (около 250п.н.) некодирующими отрезками ДНК (интронами). Промежутки между отдельными генами составляют в среднем 4.6 тыс. пар нуклеотидов. Для сравнения укажем, что гены человека содержат многие десятки и даже сотни экзонов и интронов, а межгенные участки имеют размеры от 10 тыс. пар нуклеотидов и более. Предполагают, что наличие небольшого компактного генома способствовало эволюционной устойчивости арабидопсиса, поскольку его ДНК в меньшей степени становилась мишенью для воздействия различных повреждающих агентов, в частности, для внедрения в геном вирусоподобных повторяющихся фрагментов ДНК (транспозонов).
Из других молекулярных особенностей генома арабидопсиса следует отметить обогащенность экзонов гуанином и цитозином (44% в экзонах и 32% в интронах) по сравнению с генами животных, а также присутствие дважды повторенных (дуплицированных) генов. Предполагают, что такое удвоение произошло в результате четырех одномоментных событий, заключавшихся в удвоении (повторении) части генов арабидопсиса, или слияния родственных геномов. Эти события, имевшие место 100-200 млн. лет назад, - проявление общей тенденции к полиплоидизации (кратному увеличению числа геномов в организме), характерной для геномов растений. Однако некоторые факты показывают, что у арабидопсиса удвоеннные гены неидентичны и функционируют по-разному, что может быть связано с мутациями в их регуляторных участках.
Еще одним объектом полного секвенирования ДНК стал рис [8]. Геном этого растения тоже невелик (12 хромосом, дающих в сумме 420-470 млн. п.н.), всего в 3.5 раза больше, чем у арабидопсиса. Однако, в отличие от арабидопсиса, рис имеет огромное хозяйственное значение, являясь основой питания для более чем половины человечества, поэтому в улучшении его свойств кровно заинтересованы не только миллиарды потребителей, но и многомиллионная армия людей, активно вовлеченная в весьма трудоемкий процесс его выращивания.
Отдельные исследователи приступили к изучению генома риса еще в 80-х годах прошлого столетия, но серьезного масштаба эти работы достигли лишь в 90-х. В 1991 г. в Японии была создана программа по расшифровке структуры генома риса, объединившая усилия многих исследовательских групп. В 1997 г. на базе этой программы был организован Международный проект "Геном риса" [8]. Его участники решили сконцентрировать усилия на секвенировании одного из подвидов риса (Oriza sativajaponica), в изучении которого к тому времени уже были достигнуты значительные успехи. Серьезным стимулом и, образно выражаясь, путеводной звездой для такой работы стала программа "Геном человека" [9].
В рамках этой программы прошла апробацию стратегия "похромосомного" иерархического разделения генома, которую участники международного консорциума использовали при расшифровке генома риса. Однако, если при изучении генома человека с помощью различных приемов выделяли фракции отдельных хромосом, то материал, специфичный для индивидуальных хромосом риса и их отдельных участков, получали методом лазерной микродиссекции (вырезания микроскопических объектов). На предметном стекле микроскопа, где находятся хромосомы риса, под воздействием лазерного луча выжигается все, кроме хромосомы или ее участков, намеченных для анализа. Оставшийся материал используют для клонирования и секвенирования.
Опубликованы многочисленные сообщения о результатах секвенирования отдельных фрагментов генома риса, осуществленного с высокой точностью и детальностью, характерной для иерархической технологии. Считали, что определение полной первичной структуры генома риса будет завершено к концу 2003-середине 2004 г. и результаты вместе с данными по первичной структуре генома арабидопсиса будут широко использоваться в сравнительной геномике других растений.
Однако в начале 2002 г. две исследовательские группы - одна из Китая, другая из Швейцарии и США - опубликовали результаты полного чернового (приблизительного) секвенирования генома риса, выполненного с помощью технологии тотального клонирования [10]. В отличие от поэтапного (иерархического) изучения, тотальный подход основан на одномоментном клонировании всей геномной ДНК в одном из вирусных или бактериальных векторов и получении значительного (огромного для средних и крупных геномов) количества отдельных клонов, содержащих различные отрезки ДНК. На основании анализа этих секвенированных участков и наложения друг на друга идентичных концевых участков ДНК образуется контиг - цепочка стыкованных между собой последовательностей ДНК. Общий (суммарный) контиг представляет собой первичную структуру всего генома или, по крайней мере, индивидуальной хромосомы.
В таком схематичном изложении стратегия тотального клонирования кажется несложной. На деле она встречает серьезные трудности, связанные с необходимостью получения огромного количества клонов (принято считать, что изучаемый геном или его участок должен быть перекрыт клонами, по крайней мере, 10 раз), гигантским объемом секвенирования и чрезвычайно сложной работой по стыковке клонов, требующей участия специалистов по биоинформатике. Серьезным препятствием на пути тотального клонирования служат разнообразные повторяющиеся участки ДНК, число которых, как уже упоминалось, резко возрастает по мере увеличения размера генома. Поэтому стратегию тотального секвенирования используют преимущественно при изучении геномов вирусов и микроорганизмов, хотя она и была успешно применена для исследования генома многоклеточного организма -дрозофилы.
Результаты тотального секвенирования этого генома были "наложены" на огромный массив сведений о его хромосомной, генной и молекулярной структуре, полученных за почти 100-летний период изучения дрозофилы. И все же по степени секвенированности геном дрозофилы (66% общего размера генома) значительно уступает геному арабидопсиса (92%), несмотря на достаточно близкие их размеры - 180 млн. и 125 млн. пар нуклеотидов соответственно. Поэтому недавно предложено [11] называть смешанной технологию, с помощью которой проводилось секвенирование генома дрозофилы.
Для секвенирования генома риса упомянутые выше исследовательские группы взяли два его подвида, наиболее широко культивируемые в азиатских странах, - Oriza saliva L. ssp indicaj и Oriza saliva L. sspjaponica. Результаты их исследований во многом совпадают, но во многом и различаются. Так, представители обеих групп заявили, что ими достигнуто перекрывание контигами приблизительно 92-93% генома. Показано, что около 42% генома риса представлено короткими повторами ДНК, состоящими из 20 пар нуклеотидов, и большинство подвижных ДНК-элементов (транспозонов) находится в межгенных участках. Однако сведения о размерах генома риса существенно различаются.
Для японского подвида размер генома определен равным 466 млн. пар нуклеотидов, а для индийского - 420 млн. Причина такого расхождения не ясна. Оно может быть следствием различных методических подходов в определении размеров некодирующей части геномов, то есть не отражать истинного положения дел. Но не исключено, что 15%-ное различие в размере изученных геномов действительно существует.
Второе серьезное расхождение выявилось в числе обнаруженных генов: для японского подвида - от 46022 до 55615 генов на геном, а для индийского - от 32000 до 50000. Причина такого расхождения не ясна.
Неполнота и противоречивость полученных сведений отмечена в комментариях к опубликованным статьям [10]. Здесь же высказана надежда, что пробелы в знаниях генома риса будут устранены при сопоставлении данных "чернового секвенирования" с результатами детального, иерархического секвенирования, проводимого участниками Международного проекта "Геном риса".
СРАВНИТЕЛЬНАЯ И ФУНКЦИОНАЛЬНАЯ ГЕНОМИКА РАСТЕНИЙ
Полученные обширные данные, половина из которых (результаты китайской группы) общедоступны, несомненно, открывают широкие перспективы как для изучения генома риса, так и для геномики растений в целом. Сравнение свойств геномов арабидопсиса и риса показало, что большая часть генов (до 80%), выявленных в геноме арабидопсиса, обнаружена и в геноме риса, однако приблизительно для половины генов, обнаруженных у риса, пока не удалось найти аналогов (ортологов) в геноме арабидопсиса. В то же время 98% генов, первичная структура которых установлена для других злаков, выявлены в геноме риса.
Вызывает недоумение существенное (почти в два раза) расхождение в числе генов у риса и арабидопсиса. При этом данные черновой расшифровки генома риса, полученные с помощью тотального секвенирования, практически не сопоставлены с обширными результатами изучения генома риса методом иерархического клонирования и секвенирования, то есть не осуществлено то, что сделано в отношении генома дрозофилы. Поэтому остается неясным, отражает ли различие числа генов у арабидопсиса и риса истинное положение дел или же оно объясняется различием в методических подходах.
В отличие от генома арабидопсиса, сведения о генах-двойниках в геноме риса не приведены. Не исключено, что их относительное количество может быть больше у риса, чем у арабидопсиса. В пользу такой возможности косвенно свидетельствуют данные о наличии полиплоидных форм риса. Большей ясности в этом вопросе можно ожидать после завершения Международного проекта "Геном риса" и получения детальной картины первичной структуры ДНК этого генома. Серьезные основания для такой надежды дает тот факт, что после выхода в свет работ о черновом секвенировании генома риса резко увеличилось число публикаций о структуре этого генома, в частности, появились сведения о детальном секвенировании его 1 и 4 хромосом.
Знание, хотя бы приблизительное, числа генов у растений имеет принципиальное значение для сравнительной геномики растений. Вначале считали, что поскольку по своим фенотипическим признакам все цветковые растения весьма близки друг к другу, так же близки должны быть и их геномы. И если мы изучим геном арабидопсиса, то получим сведения и о большинстве геномов других растений. Косвенным подтверждением такого допущения служат результаты секвенирования генома мыши [11], который удивительно близок к геному человека (около 30 тыс. генов, из которых различными оказалась лишь 1 тыс.).
Можно предположить, что причина различий геномов арабидопсиса и риса кроется в их принадлежности к разным классам растений - двудольным и однодольным. Чтобы прояснить этот вопрос, крайне желательно знать хотя бы черновую первичную структуру еще какого-нибудь однодольного растения. Наиболее реальным кандидатом может стать кукуруза, геном которой приблизительно равен геному человека, но все-таки значительно меньше геномов других злаков. Продовольственное значение кукурузы общеизвестно.
Огромный материал, полученный в результате секвенирования геномов арабидопсиса и риса, постепенно становится основой для широкомасштабного изучения геномов растений методами сравнительной геномики. Такие исследования имеют общебиологическое значение, так как позволяют установить главные принципы организации генома растений в целом и их отдельных хромосом, выявить общие черты структуры генов и их регуляторных участков, рассмотреть соотношение функционально активной (генной) части хромосомы и различных не кодирующих белки межгенных участков ДНК. Сравнительная генетика приобретает все большее значение и для развития функциональной геномики человека. Именно для проведения сравнительных исследований осуществлено секвенирование геномов рыбы фугу и мыши.
Не менее важно изучение отдельных генов, ответственных за синтез индивидуальных белков, определяющих конкретные функции организма. Именно в обнаружении, выделении, секвенировании и установлении функции отдельных генов состоит практическое, в первую очередь медицинское, значение программы "Геном человека". Это обстоятельство несколько лет назад отметил Дж. Уотсон, подчеркнувший, что программа "Геном человека" будет завершена лишь тогда, когда определят функции всех генов человека.
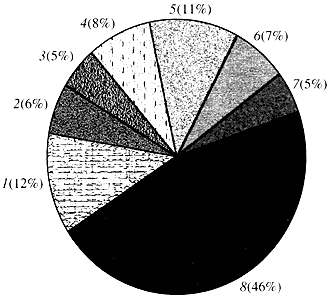
Рис. 4. Классификация по функции генов арабидопсиса
1 - гены роста, деления и синтеза ДНК; 2 - гены синтеза РНК (транскрипция); 3 - гены синтеза и модификации белков; 4 - гены развития, старения и смерти клеток; 5 - гены клеточного метаболизма и энергетического обмена; 6 - гены межклеточного взаимодействия и передачи сигнала; 7 - гены обеспечения прочих клеточных процессов; 8 - гены с неизвестной функциейЧто касается функции генов растений, то мы знаем о них менее одной десятой того, что нам известно о генах человека. Даже у арабидопсиса, геном которого по степени изученности намного превосходит геном человека, функция почти половины его генов остается неизвестной (рис. 4). Между тем у растений, кроме генов, общих с животными, имеется значительное число генов, специфичных только (или, по крайней мере, преимущественно) для них. Речь идет о генах, вовлеченных в транспорт воды и синтез клеточной стенки, отсутствующей у животных, о генах, обеспечивающих образование и функционирование хлоропластов, фотосинтез, фиксацию азота и синтез многочисленных ароматических продуктов. Этот перечень можно продолжить, но уже сейчас ясно, сколь сложная задача стоит перед функциональной геномикой растений.Полное секвенирование генома дает близкие к истинным сведения об общем количестве генов данного организма, позволяет поместить в банки данных более или менее подробные и достоверные сведения об их структуре, облегчает работу по выделению и изучению индивидуальных генов. Однако секвенирование генома отнюдь не означает установления функции всех генов.
Один из наиболее перспективных подходов функциональной геномики базируется на выявлении работающих генов, на которых идет транскрипция (считывание) мРНК. Этот подход, в том числе использующий современную технологию микрочипов, позволяет одновременно выявлять до десятков тысяч функционирующих генов. Недавно с помощью такого подхода начато изучение геномов растений. Для арабидопсиса удалось получить около 26 тыс. индивидуальных транскриптов, что резко облегчает возможность определения функции практически всех его генов [12]. У картофеля удалось выявить около 20000 тыс. работающих генов, важных для понимания как процессов роста и формирования клубня, так и процессов заболевания картофеля. Предполагается, что это знание позволит повысить устойчивость одного из важнейших пищевых продуктов к возбудителям заболеваний.
Логичным развитием функциональной геномики стала протеомика. Эта новая область науки изучает протеом, под которым обычно подразумевают полный набор белков в клетке в конкретный момент. Такой набор белков, отражающий функциональное состояние генома, все время меняется, тогда как геном остается неизменным.
Изучение белков уже давно используют для суждений об активности геномов растений. Как известно, ферменты, имеющиеся у всех растений, различаются у отдельных видов и сортов последовательностью аминокислот. Такие ферменты, с одинаковой функцией, но разной последовательностью отдельных аминокислот, называют изоферментами. У них разные физико-химические и иммунологические свойства (молекулярная масса, заряд), что можно выявить с помощью хро-матографии или электрофореза. В течение многих лет эти методы успешно использовали для изучения так называемого генетического полиморфизма, то есть различий между организмами, сортами, популяциями, видами, в частности пшеницы и родственных форм злаков. Однако в последнее время в связи с быстрым развитием методов анализа ДНК, включая секвенирование, изучение белкового полиморфизма оказалось замененным исследованием полиморфизма ДНК. Впрочем, прямое изучение спектров запасных белков (проламины, глиадины и др.), определяющих основные питательные свойства злаков, остается важным и надежным способом генетического анализа, селекции и семеноводства сельскохозяйственных растений.
Знание генов, механизмов их экспрессии и регуляции чрезвычайно важно для развития биотехнологии и получения трансгенных растений. Известно, что впечатляющие успехи в этой области вызывают неоднозначную реакцию экологической и медицинской общественности. Однако есть область биотехнологии растений, где эти страхи, если не совсем беспочвенны, то, во всяком случае, представляются малосущественными. Речь идет о создании трансгенных технических растений, не используемых в качестве пищевых продуктов. Недавно в Индии собран первый урожай трансгенного хлопка, устойчивого к ряду заболеваний [13]. Имеются сведения о введении в геном хлопка специальных генов, кодирующих пигментные белки, и получении волокон хлопка, не нуждающихся в искусственном окрашивании. Другая техническая культура, которая может оказаться объектом эффективной генной инженерии, - это лен. Его использование как альтернативы хлопку для получения текстильного сырья обсуждается в последнее время [14]. Эта проблема чрезвычайно важна для нашей страны, утратившей собственные источники хлопкового сырья.
ПЕРСПЕКТИВЫ ИЗУЧЕНИЯ ГЕНОМОВ РАСТЕНИЙ
Очевидно, что структурные исследования геномов растений будут базироваться на подходах и методах сравнительной геномики с использованием в качестве основного материала результатов расшифровки геномов арабидопсиса и риса. Существенную роль в развитии сравнительной геномики растений будут, без сомнения, играть сведения, которые рано или поздно предоставит тотальное (черновое) секвенирование геномов других растений. При этом сравнительная геномика растений будет основываться на установлении генетических взаимосвязей отдельных локусов и хромосом, относящихся к разным геномам. Речь пойдет не столько об общей геномике растений, сколько об избирательной геномике отдельных хромосомных локусов. Так, недавно было показано, что ген, ответственный за яровизацию, расположен в локусе VRn-AI хромосомы 5А гексаплоидной пшеницы и локусе Hd-6 хромосомы 3 риса.
Развитие этих исследований явится мощным толчком к идентификации, выделению и секвени-рованию многих функционально важных генов растений, в частности генов, ответственных за устойчивость к болезням, засухоустойчивость, приспособленность к различным условиям произрастания. Все шире будет использоваться функциональная геномика, основанная на массовом выявлении (скринировании) генов, функционирующих в растениях.
Можно предвидеть дальнейшее совершенствование хромосомных технологий, прежде всего метода микродиссекции. Его использование резко расширяет возможности геномных исследований, не требуя огромных затрат, как, например, тотальное секвенирование геномов. Получит дальнейшее распространение метод локализации на хромосомах растений отдельных генов с помощью гибридизации in situ. В настоящий момент его применение ограничено огромным числом повторяющихся последовательностей в геноме растений, а возможно, и особенностями структурной организации хромосом растений.
Хромосомные технологии в обозримом будущем приобретут большое значение и для эволюционной геномики растений. Эти технологии, относительно недорогие, позволяют быстро оценивать внутри- и межвидовую вариабельность, изучать сложные аллополиплоидные геномы тетраплоидной и гексаплоидной пшеницы, тритикале; анализировать эволюционные процессы на хромосомном уровне; исследовать образование синтетических геномов и введение (интрогрессия) чужеродного генетического материала; выявлять генетические взаимоотношения между индивидуальными хромосомами различных видов.
Изучение кариотипа растений с помощью классических цитогенетических методов, обогащаемых молекулярно-биологическим анализом и компьютерными технологиями, будет использоваться для характеристики генома. Это особенно важно для изучения стабильности и изменчивости кариотипа на уровне не только отдельных организмов, но и популяции, сорта и вида. Наконец, трудно представить, каким образом можно оценить число и спектры хромосомных перестроек (аберрации, мосты) без применения методов дифференциального окрашивания. Такие исследования крайне перспективны для мониторинга окружающей среды по состоянию генома растений.
В современной России вряд ли будет проводиться прямое секвенирование геномов растений. Такие работы, требующие крупных вложений, непосильны для нашей нынешней экономики. Между тем сведений о строении геномов арабидопсиса и риса, полученных мировой наукой и доступных в международных банках данных, достаточно для развития отечественной геномики растений. Можно предвидеть расширение исследований геномов растений, основанных на подходах сравнительной геномики, для решения конкретных задач селекции и растениеводства, а также изучения происхождения различных видов растений, имеющих важное хозяйственное значение.
Можно полагать, что в отечественной селекционной практике и растениеводстве будут широко использоваться такие геномные подходы, как генетическое типирование (RELF, RAPD, AFLP-анализы и т.п.), вполне доступные для нашего бюджета. Параллельно с прямыми методами определения ДНК-полиморфизма для решения проблем генетики и селекции растений будут применяться подходы, основанные на изучении белкового полиморфизма, в первую очередь запасных белков злаков. Широкое применение получат хромосомные технологии. Они относительно недороги, их развитие требует вполне умеренных вложений. В области хромосомных исследований отечественная наука не уступает мировой.
Следует подчеркнуть, что наша наука внесла заметный вклад в становление и развитие геномики растений [1, 15].
Основополагающую роль сыграл Н.И. Вавилов (1887-1943).
В молекулярной биологии и геномике растений очевиден пионерский вклад А.Н. Белозерского (1905-1972).
В области хромосомных исследований необходимо отметить работы выдающегося генетика С.Г. Навашина (1857-1930), впервые обнаружившего у растений спутниковые хромосомы и доказавшего, что можно различать отдельные хромосомы по особенностям их морфологии.
Другой классик российской науки Г.А. Левицкий (1878-1942) детально описал хромосомы ржи, пшеницы, ячменя, гороха и сахарной свеклы, ввел в науку термин "кариотип" и развил учение о нем.
Современные специалисты, опираясь на достижения мировой науки, могут внести заметный вклад в дальнейшее развитие генетики и геномики растений.
Автор выражает сердечную благодарность академику Ю.П. Алтухову за критическое обсуждение статьи и ценные советы.Работа коллектива, возглавляемого автором статьи, выполнена при поддержке Российского фонда фундаментальных исследований (гранты № 99-04-48832; 00-04-49036; 00-04-81086), Программы Президента Российской Федерации по поддержке научных школ (гранты № 00-115-97833 и НШ-1794.2003.4) и Программы Российской академии наук "Молекулярно-генетические и хромосомные маркеры в разработке современных методов селекции и семеноводства".
ЛИТЕРАТУРА
1. Зеленин А.В., Бадаева Е.Д., Муравенко О.В. Введение в геномику растений // Молекулярная биология. 2001. Т. 35. С. 339-348.
2. Pen E. Bonanza for Plant Genomics // Science. 1998. V. 282. P. 652-654.
3. Plant genomics // Proc. Natl. Acad. Sci. USA. 1998. V. 95. P. 1962-2032.
4. Картель Н.А. и др. Генетика. Энциклопедический словарь. Минск: Technologia, 1999.
5. Badaeva E.D., Friebe B., Gill B.S. 1996. Genome differentiation in Aegilops. 1. Distribution of highly repetitive DNA sequences on chromosomes of diploid species // Genome. 1996. V. 39. P. 293-306.
6. Муравенко О.В., Лемеш В.А., Саматадзе Т.Е. и др. Сравнение геномов трех близкородственных видов льна и их гибридов с использованием хромосомных и молекулярных маркеров // Генетика. 2003. Т. 39. С. 510-518.
7. Arabidopsis thaliana genome sequence // Nature. 2000. V. 408. P. 791-826.
8. Sasaki Т. The rice genome project in Japan // Proc. Natl. Acad. Sci. USA. 1998. V. 95. P. 2027, 2028.
9. Collins F.S., Morgan M., Patrikus A. The human genome project: lessons from large-scale bioligy // Science. 2003. V. 300. P. 289-290.
10. Rice Genome Sequence // Science. 2002. V. 296. P. 13, 32-39,41-46,53-56,79-100.
11. Mouse genome // Nature. 2002. V. 420. P. 456, 457. 509-590.
12. Сhen ZJ., Wang J., Lee H. et al. Genomic-wide transcriptone analysis and mechanisms of differential gene expression in Arabidopsis polyploids //Item. Polyploidy Conference. Linnean Society and Royal Botanic Garden. Kew, 2003. P. 5.
13. Yimin D., Mervis J. China takes a bumpy road from the lab to the field // Science. 2002. V. 298. P. 2317-2318.
14. Ebskamp MJ.M. Engineering flax and hemp for alternative to cotton // Trends in Biotechnology. 2002. V. 20. P. 297-298.
15. Зощук Н.В., Бадаева Е.Д., Зеленин А.В. История хромосомного анализа // Биол. мембраны. 2001. Т. 18. С. 164-172.