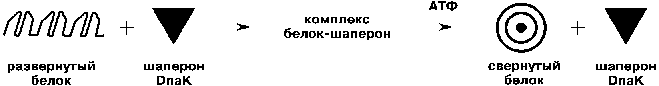№ 2, 1998 г.
№ 2, 1998 г.
№ 2, 1998 г.
№ 2, 1998 г.
© Л.Ю. Бровко, Н.Н. Угарова
Тайны и загадки "живого" света
Л.Ю. Бровко, Н.Н. Угарова
ОТ БОЙЛЯ ДО СОВРЕМЕННОСТИ
Биолюминесценция - это свет живой природы, волшебное свечение, которым можно любоваться на берегу моря, наблюдая ночью за фосфоресцирующими морскими волнами или за хороводами жучков-светлячков. Подводное царство населено множеством светящихся рыб, моллюсков, медуз и других обитателей морских глубин. Тайны происхождения "живого" света давно привлекали внимание людей. Светящиеся организмы были описаны в древнекитайских книгах еще 3000 лет назад, а также в трудах древних философов - Аристотеля и Плиния.
В 1668 г. Р.Бойль отметил нечто общее между процессами горения угля и излучения "холодного света" гниющими деревьями. В обоих случаях, как показал Бойль, свечение исчезает, если удалить воздух (т.е. кислород). Следовательно, излучение света живыми существами возникает при окислении, протекающем в клетках. Однако две главные проблемы оставались неразрешенными до второй половины XIX в.: действительно ли свет - это результат жизнедеятельности живых существ; в результате каких - физических или химических - внутриклеточных процессов появляется "холодное свечение".
Важным этапом в раскрытии тайны "живого" света стали работы французского ученого Р. Дюбуа, который выделил из жуков-светляков и из светящихся двустворчатых моллюсков две фракции. Одна из них не содержала белка и была устойчива к высокой температуре, а вторая, белковая фракция, легко теряла свои свойства при нагревании. Впоследствии термостабильную фракцию назвали люциферином, а термолабильную - люциферазой. Свечение возникало только при соединении люциферина и люциферазы и только в присутствии атмосферного кислорода, а предварительно прогретая люцифераза была не способна излучать "живой" свет. Работа Дюбуа дала толчок к поиску и изучению тех веществ в биологических системах, которые участвуют в биолюминесценции - генерировании "живого" света.
Излучение энергии в виде света присуще и чисто химическим процессам. Так, свечение, или хемилюминесценция, довольно часто возникает в реакциях окисления или при рекомбинации радикалов. Для его возникновения прежде всего необходимо, чтобы общая выделяемая энергия была больше 40 ккал/моль (это соответствует диапазону энергий видимой области спектра 400 - 700 нм). Кроме того, продуктом реакции должно быть вещество с энергией электронно-возбужденного состояния, не превышающей свободную энергию химической реакции. В этих условиях может образоваться электронно-возбужденный продукт, возвращение которого в основное состояние тоже сопровождается излучением видимого света. Свечение это довольно слабое, поскольку квантовый выход электронно-возбужденного продукта невысок.
В отличие от хемилюминесцентных, биолюминесцентные, живые, системы светятся необычайно ярко. Это объясняется тем, что почти каждая молекула продукта реакции образуется в электронно-возбужденном состоянии. Биолюминесцентные системы имеют ряд существенных особенностей. Во-первых, реакции с выделением света протекают в водных растворах при значениях рН, близких к нейтральным, когда многие хемилюминесцентные реакции неэффективны. Во-вторых, несмотря на водную среду, квантовый выход биолюминесцентных реакций очень высок (0.1 - 1.0). Естественно, что столь значительная эффективность процесса достигается за счет участия высокоспецифичных белковых биокатализаторов (ферментов).
В настоящее время расшифрованы структуры основных компонентов - люциферинов и люцифераз - из наиболее известных биолюминесцентных систем. Оказалось, что люциферины - это сложные органические молекулы разнообразной химической структуры. Примечательно, что у светящихся организмов, принадлежащих таксонам высокого ранга (начиная с класса), структура люциферинов сильно различается, лишь у представителей немногих классов она сходна. Но видовая специфичность в строении люциферинов отсутствует. Иными словами, например у бактерий (царство прокариот), медуз (жизненная форма животных типа книдарий), насекомых (класс) структура этих молекул имеет мало сходства, а у всех изученных видов светляков содержится один и тот же люциферин.
Основную роль в биолюминесцентных системах играют ферменты люциферазы. Это те матрицы, на которых протекает высокоспецифичное окисление молекулы люциферина в электронно-возбужденный продукт. С другой стороны, люциферазы управляют этим процессом, ускоряя его в тысячи раз и направляя по пути, ведущему к выделению света. В отсутствие люцифераз люциферины тоже можно окислить, но в этом случае скорость процесса будет мала, а интенсивность свечения в сотни тысяч раз слабее, чем при ферментативном катализе.
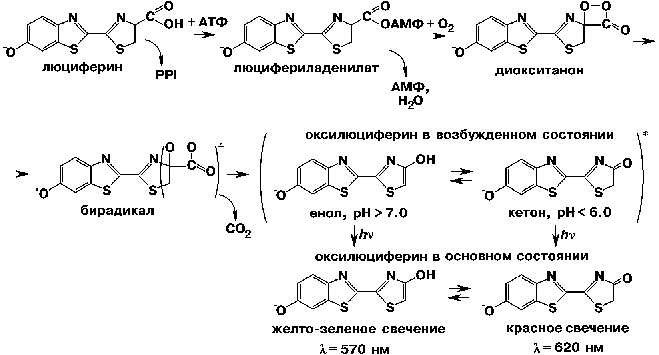
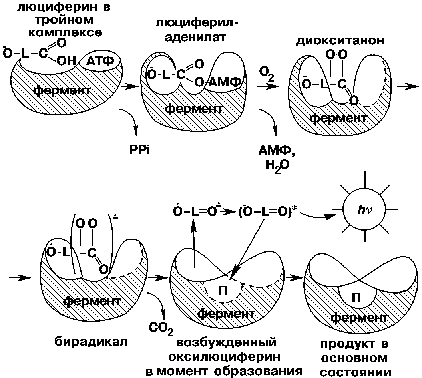
Химические механизмы окисления люциферинов у разных классов светящихся организмов существенно различаются. Для примера приведем биолюминесцентную систему жуков-светляков. На первой стадии фермент связывается со своими субстратами - люциферином и аденозин-5'-трифосфатом (АТФ). В таком тройном фермент-субстратном комплексе люциферин ковалентно взаимодействует с АТФ, образуются смешанный ангидрид карбоновой и фосфорной кислот, люцифериладенилат и пирофосфат. Люцифериладенилат в комплексе с ферментом через ряд промежуточных стадий окисляется кислородом воздуха, превращаясь в циклический пероксид, диокситанон. Эта молекула имеет замечательную особенность: одна ее часть - легко окисляемая гетероциклическая структура (люциферил) - обладает низким ионизационным потенциалом, а другая (пероксид) - высоким сродством к электрону. За счет внутримолекулярного переноса электрона от фенолятной группы к пероксидной образуется резонансная структура. Разрыв О - О-связи вызывает декарбоксилирование диокситанона и возникновение бирадикала - анион-радикала кетона и катион-радикала фенолята. Конечный продукт реакции - оксилюциферин в синглетном электронно-возбужденном состоянии - образуется в результате внутримолекулярной аннигиляции этих радикалов. Когда оксилюциферин переходит в свое основное состояние, наблюдается желто-зеленое свечение (550 - 570 нм).
Несмотря на значительный прогресс, достигнутый в последние годы в изучении биолюминесценции, многие тайны еще остаются нераскрытыми. В течение ряда лет мы детально изучаем структуру и механизм действия люциферазы светляков с целью выяснить роль белковой глобулы фермента в процессе высокоэффективной трансформации химической энергии в световую. Свое внимание мы сосредоточили на исследовании механизма излучения; кинетических характеристик ферментативной реакции; на выяснении роли белка как матрицы, создающей оптимальную микросреду.
Все это требовало больших количеств люциферазы, которую до недавнего времени выделяли из светляков. Чтобы получить 1 мг чистого фермента, необходимо огромное число этих жуков: 800 - 1000 особей. Но в последние годы был выделен ген люциферазы и включен в клетки кишечной палочки (Esсherichia coli). В результате появилась возможность получать практически неограниченные количества люциферазы светляков в лабораторных условиях. Правда, оставалась существенная проблема - оптимизировать биосинтез активной люциферзы в клетках Е.coli.
БИОСИНТЕЗ ЛЮЦИФЕРАЗЫ СВЕТЛЯКОВ В БАКТЕРИАЛЬНЫХ КЛЕТКАХ
Успехи современной генной инженерии позволяют синтезировать в бактериальных клетках чужеродные им белки. Эта технология широко используется в научных исследованиях и для производства практически важных продуктов. Известно, что пространственная структура белка определяет его функциональную активность. Сворачивание белковой цепи, т.е. образование конечной пространственной структуры вновь синтезированного белка в клетках родительского организма, - процесс, во многом еще загадочный. Рассуждая логически, можно предположить, что организм каким-то образом "знает", какой должна быть пространственная конформация белка, чтобы он выполнял требуемые от него функции. Когда же мы искусственно переносим процесс синтеза белка, животного или растительного, в чужеродное окружение бактериальной клетки, обретение белком нужной конформации становится особенно важным. Но нередко синтезированный в микробной клетке белок не обладает правильной пространственной структурой и, следовательно, желаемыми свойствами, хотя его химический состав не отличается от природного.
При исследовании процесса биосинтеза люциферазы светляков в клетках E.coli мы обнаружили, что на начальной стадии белок синтезируется в каталитически неактивной форме. Его превращение в активный фермент требует значительного времени - до 50 - 70 ч. Не будучи самопроизвольным, этот процесс управляется особыми белками - шаперонами. Их функции весьма разнообразны, но основная их задача - контроль за правильностью сворачивания синтезируемых в клетке белков. И этот контроль шапероны осуществляют различными путями. Одни шапероны связываются с "неправильно" свернутыми или агрегированными белками, заставляя их развернуться и после этого вновь свернуться, но уже правильно. Другие шапероны образуют комплексы с синтезированным белком, который еще не приобрел конечной активной формы. Когда такой комплекс разрушается, высвобождается каталитически активный фермент.
Мы доказали, что именно такой механизм реализуется при биосинтезе люциферазы светляков в клетках E.coli благодаря наличию в них шаперона DnaK. В дефектных штаммах E.coli, где такой шаперон не синтезируется, образовавшийся белок люциферазы не превращается в каталитически активный фермент. Шапероны называют белками теплового шока, поскольку их синтез в клетках инициируется такими стрессовыми воздействиями, как резкое повышение температуры или рН среды. Для функционирования эти белки требуют участия АТФ (как источника необходимой энергии). Поэтому, когда мы подвергали клетки Е.сoli кратковременному тепловому шоку да еще вводили в них дополнительные количества АТФ, нам удавалось не только увеличить активность синтезированной люциферазы, но и в несколько раз ускорить ее сворачивание, а стало быть, заметно оптимизировать биосинтез фермента.
Аналоги шаперонов DnaK имеются не только в бактериальных, но и в животных клетках. Можно предположить, что и в организме самих светляков сворачивание белка люциферазы идет таким же образом.
ЛЮЦИФЕРАЗА - ФЕРМЕНТ-"САМОУБИЙЦА"?
В природе в сумеречное время светляки ярко светятся не один час подряд, подмигивая своими "лампочками" с частотой в несколько секунд. В пробирке же при смешении всех необходимых компонентов биолюминесцентной реакции видна яркая, но короткая вспышка света. Чем выше концентрации люциферина, АТФ и люциферазы в реакционной смеси, тем вспышка ярче. А затем наблюдается лишь слабое остаточное свечение, которое медленно затухает в течение нескольких часов. Само собой напрашивается объяснение, что быстрое затухание свечения связано с расходованием реагирующих веществ. Но в действительности после вспышки их концентрации в реакционной смеси достаточно высоки. Почему же реакция как бы останавливается, не доходя до конца?
Чтобы разрешить эту загадку, мы проанализировали (с помощью компьютерных программ) кинетику биолюминесцентной реакции при разном соотношении концентраций люциферазы, люциферина и АТФ. Выяснилось, что фермент инактивирует, теряет каталитическую активность, на всех стадиях ферментативного окисления люциферина. Постепенно инактивируют также и комплексы фермента с субстратами, промежуточным и конечным продуктами. Быстрее всего - в десятки раз по сравнению со свободным ферментом - теряет активность комплекс люциферазы с продуктом. Кроме того, этот комплекс настолько прочен, что при избытке субстратов после первого цикла реакции большая доля фермента так и остается полностью связанной, а значит, неактивной в последующих каталитических циклах.
Возникает вопрос, почему же этого не происходит в организме самих светляков? По-видимому, в их клетках присутствуют специальные вещества, которые защищают фермент от инактивации и способствуют разрушению комплекса фермент - продукт, т.е. регенерации активного биокатализатора. Действительно, мы обнаружили, что некоторые поверхностно-активные и природные вещества обладают такой способностью. Еще несколько лет назад было показано, что если добавить коэнзим А (один из природных коферментов, участвующий в реакциях окисления, синтеза жирных кислот и т.д.) к уже "потухшей" реакционной смеси, то возникает повторная вспышка света. Особенно интересные эффекты мы наблюдали, когда биолюминесцентную реакцию проводили не просто в водном растворе, а в мембраноподобном окружении: в мицеллах или липосомах. В этом случае яркость свечения не только возрастала в десятки раз по сравнению с водным раствором, но и не ослабевала в течение десятков минут. Проще говоря, вместо кратковременной вспышки мы получали яркую "лампочку". Эти эксперименты с очевидностью показывают, что в организме светляка люцифераза находится не в водном окружении, а в особых внутриклеточных мембранных структурах. Они-то и обеспечивают наиболее эффективное функционирование фермента и почти 100%-е преобразование энергии ферментативной реакции в световую.
ПОЧЕМУ СВЕЧЕНИЕ БЫВАЕТ РАЗНЫМ?
В природе жуки-светляки отличаются размерами и формой, а цвет излучаемого света у разных видов светляков варьируется от зеленого до оранжевого, т.е. максимум в спектре биолюминесценции изменяется от 490 до 590 нм. Ямайские жуки-щелкуны имеют даже несколько световых органов, испускающих свет неодинакового цвета. В то же время химический механизм биолюминесцентной реакции, структуры исходного люциферина и продукта его окисления - оксилюциферина, излучающего свет, идентичны у всех известных видов жуков-светляков. Таким образом, различия в спектрах биолюминесценции могут быть обусловлены только вариациями в структуре люцифераз, в активном центре которых и рождается электронно-возбужденный оксилюциферин. Методами генной инженерии определены химические структуры люцифераз многих видов светляков и установлено, что их аминокислотные последовательности очень сходны, но не идентичны. С помощью компьютерного анализа первичной структуры белковых цепей всех известных люцифераз светляков и жуков-щелкунов мы выяснили, что цвет биолюминесценции зависит от физико-химических свойств фрагмента, который включает аминокислотные последовательности 223 - 247. Даже точечные замены аминокислот в нем могут приводить к изменению цвета биолюминесценции. Следовательно, вблизи этого фрагмента и находится тот участок белка, который образует микроокружение излучателя - электронно-возбужденного продукта реакции, оксилюциферина.
Чтобы понять, каким образом белковое микроокружение может влиять на спектры люминесценции оксилюциферина, рассмотрим несколько подробнее физико-химический механизм взаимодействия излучателя с окружающими его молекулами растворителя или белковыми фрагментами. Это влияние может быть как специфическим, так и неспецифическим. В первом случае излучатель химически взаимодействует с микроокружением, например, образуя водородные связи или комплексы с переносом заряда. При этом изменяется электронная структура излучателя, а в результате - и спектр люминесценции, причем сильно и дискретно. Именно за счет такого специфического механизма сдвигается спектр биолюминесценции для люциферин - люциферазной системы светляков in vitro при уменьшении рН раствора. Если значения рН среды оптимальны (7.5 - 7.8), то оксилюциферин находится в форме енола и спектр биолюминесценции имеет максимум в диапазоне 550 - 570 нм, т.е. в желто-зеленой области. При уменьшении рН ниже 6.0 протонируются некие группы белка в субстратсвязывающем центре, за счет чего меняется ионное состояние оксилюциферина. Он переходит в форму кетона, и максимум спектра смещается в более длинноволновую область - 620 нм. В среде с промежуточными значениями рН присутствуют обе формы излучателя - енольная и кетонная, и спектр люминесценции оказывается бимодальным.
С другой стороны, микроокружение излучателя (будь то в растворе или в белковом окружении) можно рассматривать как некую непрерывную диэлектрическую среду, которая оказывает неспецифическое влияние на спектр люминесценции. Энергия, выделяемая в виде света при переходе возбужденной молекулы в основное состояние, зависит от разности энергий этих двух состояний. Чем больше эта разность, тем сильнее максимум спектра люминесценции сдвинут в коротковолновую область. Уровни энергии в свою очередь зависят как от электронных структур молекулы оксилюциферина в основном и возбужденном состоянии, так и от энергии взаимодействия излучателя с растворителем, т.е. энергии сольватации. Чем она выше, тем ниже уровень энергии возбужденной молекулы, тем меньше энергия, выделяемая в виде света, и тем больше максимум излучения сдвинут в длинноволновую область при неизменной форме спектра.
Процесс сольватации молекулы излучателя зависит не только от величины энергии его взаимодействия с растворителем, но и от динамических характеристик последнего, другими словами, от скорости переориентации его молекул при контакте с молекулой излучателя. Дело в том, что при переходе оксилюциферина в электронно-возбужденное состояние мгновенно происходит перестройка его электронной структуры (за 10-15 с), резко увеличивается дипольный момент молекулы. В начальный момент возбуждения оксилюциферин находится в "неравновесном" состоянии по отношению к молекулам среды, и лишь после переориентации диполей растворителя в результате движения всей его молекулы, а не только ее электронов, энергия возбужденного состояния излучателя уменьшается. Если за время жизни молекулы возбужденного оксилюциферина (10-9 - 10-8 с) окружающие его молекулы растворителя не успевают переориентироваться, то различие энергий возбужденного и основного состояний будет максимальным, а излучение - наиболее коротковолновым. В том случае, когда молекулы растворителя успевают переориентироваться, возбужденный оксилюциферин переходит в равновесное со средой состояние, и его энергия уменьшается на величину энергии сольватации, а спектр люминесценции сдвигается в длинноволновую область тем сильнее, чем выше поляризуемость и подвижность молекул среды. При возвращении же электронно-возбужденной молекулы в основное состояние наблюдается обратный процесс. Таким образом, стабилизация основного и возбужденного состояний излучателя зависит от времени, поляризуемости и вязкости растворителя.
А как влияет на цвет биолюминесценции белковое микроокружение? Если возбужденный оксилюциферин находится в сильно неполярном микроокружении, он не способен эффективно взаимодействовать с люциферазой. Тогда различие энергий возбужденного и основного состояний излучателя будет максимальным, а излучение - наиболее коротковолновым, т.е. в сине-зеленой области спектра. К такому же результату приводит и белковое микроокружение с высокой жесткостью белкового фрагмента и низкой поляризуемостью аминокислотных остатков (хотя оно и способно эффективно взаимодействовать с излучателем). В этом случае за время жизни возбужденной молекулы оксилюциферина они не успевают переориентироваться, и излучение происходит с более высокого энергетического уровня. Но если само окружение достаточно полярно, высоки подвижность белкового фрагмента и поляризуемость аминокислотных остатков в микроокружении оксилюциферина, то последние успевают перестроиться, и энергия излучателя уменьшается. Чем сильнее его взаимодействие с белком и чем выше подвижность белкового фрагмента в активном центре фермента, тем более максимум спектра биолюминесценции сдвигается в длинноволновую, оранжево-красную, область.
Интересные дополнительные сведения о роли микроокружения излучателя и его поведении в активном центре люциферазы мы получили при изучении флуоресцентных свойств модельной системы - комплекса фермент - оксилюциферин. Флуоресценция этого комплекса являет собой полный аналог стадии излучения света в люциферин - люциферазной системе.
Известно, что каждый элементарный акт флуоресценции - это излучение плоско-поляризованного света. В невязком растворе возбужденные низкомолекулярные частицы, например оксилюциферин, обладают практически свободным вращением, и поэтому поляризация флуоресценции не регистрируется. Иное дело, если в образовании комплекса участвуют крупные частицы, например молекулы белка. Тогда скорость углового вращения излучателя значительно уменьшается, и регистрируемая степень поляризации испускаемого света будет тем выше, чем больше молекулярная масса комплекса и чем он прочнее.
Изучаемый нами комплекс люцифераза - оксилюциферин настолько прочен, что его легко выделить из реакционной среды с помощью хроматографии. Однако мы не наблюдали никаких различий в степени поляризации света, испускаемого оксилюциферином в свободном состоянии и в комплексе с люциферазой. Это может быть связано, на наш взгляд, с высокой подвижностью и свободным вращением возбужденного оксилюциферина в составе комплекса с белком. Зависимость стационарных спектров флуоресценции от pH среды и кинетика затухания процесса при импульсном режиме возбуждения указывают на то, что свойства микроокружения электронно-возбужденного оксилюциферина, когда он находится в комплексе с белком, близки к свойствам растворителя с высокой диэлектрической проницаемостью, например воды. Но ведь это противоречит другим экспериментальным фактам, которые свидетельствуют, что участок белка, связывающий оксилюциферин, гидрофобен, а не гидрофилен, как вода.
Снять противоречие позволит, вероятно, предложенная нами гипотеза "диссоциативного" механизма излучения. В процессе окисления люциферина, связанного в комплекс с ферментом, образуется, как уже упоминалось, молекула оксилюциферина в электронно-возбужденном состоянии. Энергия этой частицы столь велика, а ее окружение термодинамически столь невыгодно, что комплекс белок - продукт разрушается почти мгновенно. Переход возбужденного излучателя в основное состояние, который сопровождается свечением, происходит в околобелковом пространстве в окружении молекул воды, структурированных аминокислотными остатками. Затем оксилюциферин, будучи уже в основном состоянии, вновь связывается с люциферазой, образуя прочный комплекс. Предложенная гипотеза объясняет и зависимость цвета биолюминесценции от структуры белка, и "водоподобное" окружение электронно-возбужденного оксилюциферина, и его свободное вращение в момент излучения света.
Таким образом, кинетические и физико-химические методы исследования механизма биосинтеза и функционирования люциферазы светляков помогли нам проникнуть в некоторые тайны возникновения "живого" света. Возможно, в будущем удастся создать искусственные светящиеся биосистемы с коэффициентом полезного действия, близким к 100%. Но уже и сейчас биолюминесценция находит практическое применение. В последние годы на ее основе создан ряд чувствительнейших методов анализа широкого круга веществ. Это экспресс-методы контроля микробных загрязнений в пищевых продуктах и напитках, быстрые методы иммуноферментного анализа, целевой подбор антибиотиков, иммуномодуляторов, экспресс-диагностика инфаркта миокарда и многие другие. В природе же "живой" свет по-прежнему нужен светящимся существам: одним для привлечения добычи, другим - для защиты от врагов, третьи с его помощью отыскивают партнера, чтобы продолжить свой род.
|
|