№2, 2005 г. №2, 2005 г.
№2, 2005 г. №2, 2005 г.© Бородин П.М., Д'Андрейа П.С., Баррейро Гомес С.К.
Естественная история зверя пунаре
в 8 1/2 главахП.М. Бородин, П.С. Д'Андрейа, С.К. Баррейро Гомес
Павел Михайлович Бородин, д.б.н., зав. лаб. рекомбинационного и сегрегационного анализа
Ин-та цитологии и генетики СО РАН, профессор кафедры цитологии и генетики Новосибирского государственного университета.Пауло Серджио Д'Андрейа, доктор философии (экология), зав. лаб. биологического контроля шистосомоза
Ин-та тропической медицины им.Освальдо Круза (Рио-де-Жанейро, Бразилия).Симона Кристина Баррейро Гомес, стажер той же лаборатории.
Глава 1. Кто такой зверь пунаре и как он попал в вжную Америку
Из всех невиданных зверей, которых, как известно, в Бразилии такое изобилие, пунаре (он же рабуду, он же трихомис, он же щетинистая крыса), конечно, не самый невиданный. Он относится к роду трихомисов из семейства щетинистых крыс из подотряда кавиоморфных грызунов (рис.1). Хотя грызуны ли кавиоморфные грызуны, это для некоторых еще не очевидно. По крайней мере по поводу самого главного кавиоморфного грызуна - кавии (она же морская, а точнее, заморская свинка) в научной литературе недавно шли бурные дебаты.

Рис. 1. Самка пунаре с детенышами. Фото С.Р.Бонвицино
В журнале "Nature" в 1991 г. была опубликована статья под названием "А грызун ли морская свинка?". В ней на основе анализа последовательности аминокислот в некоторых белках утверждалось, что морская свинка отделилась от общего ствола млекопитающих задолго до того, как приматы и парнокопытные разошлись на пути эволюции с мышеобразными грызунами, а потому, скорее всего, не грызун. Через два года в "Proc. Natl. Acad. Sci. USA" появилась другая статья, авторы которой, опираясь уже на данные о последовательности нуклеотидов, доказывали, что морская свинка все-таки грызун [2]. Еще через три года опять же в "Nature" вышла статья третьей группы авторов, которую они назвали прямо и недвусмысленно "Морская свинка - не грызун". дискуссия на этом, однако не закончилась, а пошла по второму кругу. В 1997 г. опять публикуется статья других исследователей под почти тем же самым названием, что и самая первая статья, - "А грызуны ли морские свинки?". Все эти споры вызваны тем, что ветвления филогенетического древа грызунов произошли очень давно, гораздо раньше, чем выделились некоторые другие отряды млекопитающих (например, хищные и копытные).
Филогенетическое древо грызунов по современным данным выглядит примерно так. Ствол его образовался очень давно, более 90 млн лет назад. На нем выделились две основные ветви: одна к белкам и соням, а другая ко всем остальным грызунам. Затем эта последняя ветвь дала начало двум новым: одна к кавиоморфам и дикобразам, а другая ко всем остальным грызунам. И наконец, разделились ветви кавиоморф и дикобразов, общий предок которых жил около 50 млн лет назад в Африке. От нее уже откололись и ушли в долгое плавание Австралия с Индией и Антарктидой в одну сторону, а вжная Америка - в другую.
Тогда в вжной Америке не было никаких грызунов, были лишь неполнозубые (броненосцы, муравьеды и ленивцы) и отдельный, ныне вымерший, отряд копытных. Грызуны и приматы загадочным образом проникли из Африки в вжную Америку гораздо позже - около 35 млн лет назад. Как им это удалось, каким образом они смогли преодолеть Атлантический океан, не знает никто. Наиболее популярная гипотеза - с камешка на камешек (stepping-stone). Предполагается, что между Африкой и вжной Америкой существовала цепь островов, по которой приматы и кавиоморфы перебрались в вжную Америку. И тем, и другим Новый Свет понравился. Они нашли там множество разнообразных экологических ниш, с удовольствием их заселили и дали начало фантастическому разнообразию новых семейств, родов и видов. И вид трихомис (он же рабуду, он же пунаре) - один из них. Только вот один ли?
На этот вопрос мы и попытались найти ответ. Одного из нас (П.М.Бородина) интересовал сам механизм образования новых видов. Как из одного вида получается несколько? Где та грань, за которой отдельные популяции одного вида становятся разными видами? Что определяет эту грань, и как это что-то возникает в эволюции? Бородин искал ответы на вопросы в Сибири на обыкновенной землеройке [1], в японии на мускусной землеройке [2] и, наконец, в Бразилии на пунаре. П.Д'Андрейа и С.Гомес проблема интересовала в экологическом и эпидемиологическом аспектах. Дело в том, что зверь пунаре - промежуточный хозяин очень опасного паразита, трипаносомы крузи, вызывающего у людей болезнь Чагаса. Трипаносома живет в кишечнике летучих клопов-кровососов, которые имеют неприятную привычку пить кровь спящего человека и при этом опорожнять кишечник там же, где и пьют. То место, откуда клоп попил крови, начинает чесаться. В расчесы попадает трипаносома из экскрементов клопа. Затем она расселяется по организму и медленно, но верно поражает гладкую мускулатуру пищеварительного тракта и сердца. Инкубационный период болезни Чагаса длится от 10 до 20 лет.
Глава 2. Где ловили зверя пунаре
Мы попали в вжную Америку через 30 млн лет после прибытия туда кавиоморфных грызунов и обезьян Нового Света, и нам там тоже очень понравилось. Наша первая совместная экспедиция работала на территории фазенды Сертан де Формозо, что находится в самом центре Бразилии на границе штатов Гояйс, Баийя и Минас Жерайс. Все пунаре, попавшиеся здесь в ловушки, имели кариотип из 28 хромосом. Это нас несколько удивило, потому что из литературы известно, что стандартный кариотип пунаре состоит из 30 хромосом. Но поскольку по всем морфологическим показателям наши звери походили на нормальных пунаре, а один из нас по своему опыту работы с землеройками знал, что популяции одного и того же вида могут значительно отличаться по числу хромосом [1, 2], удивление было несильным. Мы взяли с собой несколько живых пунаре с этой фазенды и поселили их в виварий Института тропической медицины в Рио-де-Жанейро. Так была основана лабораторная колония Сертан де Формозо.
Следующая наша экспедиция работала на северо-востоке Бразилии в штате Пиауи. Там на зеленых склонах Сьерра де Капибара, окруженных мертвыми марсианскими лесами каатинги, расположен крохотный городок под названием Коронел (полковник) Жозе Диас. Чем был знаменит этот полковник, никто в городе не знает. Но дело не в полковнике, а в том, что неподалеку от городка в огороде у гостеприимной доны Марии мы поймали пунаре со стандартными кариотипами - 30 хромосом. Их мы тоже вывезли живыми в виварий института. А поскольку огород доны Марии назывался фазенда Фелисидадже (счастье), то и колония из ее огорода получила то же название.
В третий раз мы встретились с пунаре в месте, где их вовсе не ожидали увидеть, - в болотах Пантанала на юго-западе Бразилии. До сих пор наш зверек встречался в сухих каменистых местах и довольно высоко над уровнем моря. Но здесь на фазенде Алегрия (радость) - ниже уровня моря и в гигантском, никогда не пересыхающем болоте по соседству с крокодилами и пираньями - он тоже жил в радости, и по всем признакам это был все тот же пунаре. Правда, хромосом у него было 34. Этих зверей мы тоже привезли в виварий, и от них пошла колония под названием фазенда Алегрия.
После этого один из нас покинул Новый Свет и вернулся в Сибирь к своим старосветским землеройкам, а двое других занялись скрещиванием пунаре из всех трех колоний друг с другом. Зверь этот размножается медленно, зрелости достигает в полгода, беременность у него продолжается целых три месяца, поэтому скрещивания шли небыстро.
Глава 3. Как гибриды пунаре с пунаре соблюдали правило Холдейна
Когда через два года Бородин ненадолго вернулся в Бразилию, гибридов было уже достаточно. Выглядели они точно так же, как и их родители. Это неудивительно, поскольку родители были внешне неотличимы друг от друга. Итак, представители разных популяций пунаре скрещиваются друг с другом и дают жизнеспособное потомство. Следовательно зоологи правы, относя их к одному виду? Пока подождем с таким выводом. Мы не только получили гибридов, но и попытались их размножить, скрещивая гибридов друг другом и с представителями родительских популяций. И тут у нас возникли проблемы.
Самки-гибриды нормально скрещивались с родительскими формами и давали потомство, но гибридные самцы оказались бесплодными. Вскрытие показало, что их гонады сильно редуцированы: они были втрое меньше, чем у нормальных пунаре. Гистологический анализ обнаружил, что сперматогенез у них начинается нормально, а потом внезапно останавливается в самом начале мейоза - процесса, который ведет к образованию гамет. В гонадах гибридов между разными популяциями пунаре мы не обнаружили ни одного зрелого сперматозоида. При этом самки-гибриды были вполне плодовиты.
Здесь самое время вспомнить о правиле Холдейна. Его сформулировал в начале XX в. один из отцов-основателей синтетической теории эволюции Дж.Б.С.Холдейн (рис.2). Это одно из самых строгих и самых непонятных правил в эволюционной биологии. Строгое в том смысле, что практически не имеет исключений, а непонятное потому, что никто не понимает, почему и как оно выполняется. Правило Холдейна звучит так: "Если при гибридизации видов или популяций один пол отсутствует или стерилен, то это гетерогаметный пол", т.е. пол, у которого разные половые хромосомы. У млекопитающих и двукрылых насекомых - это самцы, имеющие хромосомы X и Y, у птиц и бабочек - это самки с половыми хромосомами W и Z. Если у гибридов один пол стерилен, то у млекопитающих и двукрылых - это всегда самцы, а у птиц и бабочек - всегда самки.
Рис. 2. Дж.Б.С.Холдейн на митинге на Трафальгарской площади. Итак, гибриды пунаре в очередной раз подтвердили справедливость правила Холдейна, от чего оно, правда, не стало понятнее. Почему гетерогаметность (гетерозиготность по половым хромосомам) приводит к стерильности? К этому вопросу мы еще вернемся. А пока обратим внимание на то, что наши стерильные самцы-гибриды были гетерозиготны не только по половым хромосомам, но и по некоторым аутосомам (неполовым хромосомам).
Глава 4. Как пунаре накапливали хромосомные перестройки
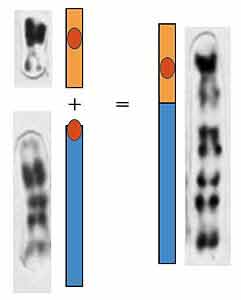
Пунаре из центра Бразилии имели 28 хромосом, с северо-востока - 30, с юго-запада - 34. Надо думать, что различия в кариотипах между популяциями образовались в ходе эволюции благодаря возникновению и локальному распространению определенных хромосомных перестроек. Видимо, из 34-хромосомного кариотипа за счет серии перестроек возник 30-хромосомный, а от него произошел 28-хромосомный благодаря тому, что две хромосомы (одна большая и одна маленькая) слились и образовали одну очень большую хромосому (рис.3).
Рис. 3. —линная хромосома у 28-хромосомных пунаре.
Произошла за счет слияния двух хромосом,
характерных для 30-хромосомных пунаре.Гибриды между носителями таких разных кариотипов оказываются гетерозиготами по тем хромосомным перестройкам, которые их отличают. Принято считать, что такая гетерозиготность ведет к стерильности из-за трудностей в спаривании и расхождении в мейозе. Это верно для определенного класса перестроек. Но в него уж точно не должны входить те, которые отличают особей одного вида или близкие виды. Почему?
Потому что каждая из этих перестроек когда-то и у кого-то возникла и была передана одному или нескольким потомкам. Ее первые носители неизбежно были гетерозиготами, потому что хромосомные перестройки возникают очень редко и абсолютно невероятно, чтобы одно и то же изменение возникло одновременно у обоих родителей. Самые первые носители перестройки не могли быть стерильны. Ведь если бы они были стерильны, то не смогли бы передать ее своим потомкам, и она не распространилась бы в популяции. Иначе говоря, если допустить, что 28-хромосомные пунаре произошли от 30-хромосомных, пройдя при этом стадию гетерозиготности по хромосомному слиянию (а иного логического пути у нас нет), то нельзя утверждать, что их гибриды стерильны из-за гетерозиготности по этому слиянию. Заметьте, что все гибриды (и самцы, и самки) гетерозиготны по слиянию, а стерильны только самцы. Итак, простая логика показывает, что стерильность самцов-гибридов не объясняется гетерозиготностью по хромосомным перестройкам.
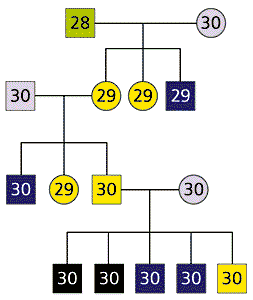
Рис. 4. Фрагмент родословной гибридов пунаре. Квадратами обозначены самцы, кружками - самки.
Зеленым - представители 28-хромосомной популяции,
розовым - 30-хромосомной.Желтым - фертильные гибриды,
синим - стерильные гибриды,
черным - еще более стерильные гибриды.Указано число хромосом у каждой особи.
Гипотеза о хромосомной стерильности отвергается и результатами скрещивания фертильных самок гибридов с самцами из родительских популяций (рис.4). При скрещивании 30-хромосомной гибридной самки с 30-хромосомным самцом все пять потомков были самцами и все они имели 30 хромосом. Среди них один был фертильным, два - такими же стерильными, как самцы первого поколения, а два - еще более стерильными. На законный вопрос читателя, как можно быть стерильней стерильного, мы ответим чуть позже. А сейчас только подчеркнем, что восстановление нормального кариотипа не привело к восстановлению фертильности у большинства самцов потомков гибридной самки. Этот факт еще раз подтверждает, что гетерозиготность по хромосомным перестройкам, отличающим популяции, не служит причиной стерильности гибридных самцов.
Так почему они все-таки стерильны? —авайте повнимательней посмотрим на потомков от скрещивания гибридной самки с самцом из родительской популяции. Вернее, не на самих потомков, а на то, что происходило в их гонадах.
Интерлюдия. Спаривание хромосом и его контролеры
Перефразируя Льва Толстого, можно сказать у всех фертильных зверей мейоз идет в принципе одинаково, а у всех стерильных он нарушается по-разному. Разберемся сначала с фертильными самцами. Мейоз начинается с того, что гомологичные (парные) хромосомы разыскивают друг друга. Как они это делают, до сих пор непонятно. Причем непонятно не только для пунаре, но и для всех живых организмов, которые размножаются половым путем. Настолько непонятно, что это заслуживает отдельного разговора [3].
Мы до сих пор не знаем, как мейотические хромосомы находят друг друга. Хотя в гипотезах недостатка нет. Многие согласны с тем, что хромосомы начинают сближаться на стадии лептотены: их концы, скользя по ядерной мембране, собираются в одной точке, и формируется структура, названная букетом. В ней гомологичные хромосомы оказываются поблизости друг от друга и приступают к взаимному опознанию. Ему посвящена следующая стадия мейоза - зиготена, которая, как предполагают некоторые специалисты, идет в два этапа: сначала приблизительное, а затем точное.
Приблизительное опознание может происходить по принципу штрих-кодов. Известно, что хромосомы представляют собой комплекс ДНК с белками. Их набор и характер связывания с ДНК во многом определяется последовательностью нуклеотидов. Поэтому каждая хромосома характеризуется индивидуальным, только для нее специфичным распределением белков - штрих-кодом. Поскольку у гомологичных хромосом последовательности ДНК в основном сходны, они должны иметь сходные штрих-коды. Таким образом, грубое распознавание может быть достигнуто простым их совмещением.
Тонкое опознание начинается с того, что в ДНК мейотических хромосом возникают множественные двунитевые разрывы. Свободные концы разорванных ДНК привлекают на себя специфические ферменты, с помощью которых они внедряются в неповрежденные районы ДНК других хромосом (не спрашивайте нас, откуда они знают, что это - ДНК других хромосом, а не их собственной, ответа не знает никто), и пытаются там отыскать комплементарные участки определенной протяженности. Найдя такой участок, внедрившаяся нить ДНК с ним спаривается. Тонкое опознание заканчивается, когда количество связок между ДНК пары гомологичных хромосом достигает критического уровня. Клетка вступает в стадию пахитены. Связи между гомологами фиксируются определенными белками, и хромосомы остаются в таком спаренном состоянии до вступления в следующую фазу мейоза - диплотену.
Вскоре после нее у всех самцов млекопитающих, включая и человека, и пунаре, следуют два цикла клеточного деления. После второго деления продукты мейоза окончательно превращаются в зрелые сперматозоиды, готовые для оплодотворения (рис.5).
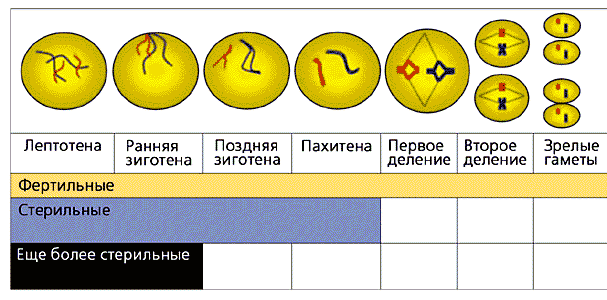
Рис. 5. Стадии мейоза млекопитающих.
Показаны только две пары гомологичных хромосом.
У фертильных пунаре мейоз доходит до конца,
у стерильных - останавливается на пахитене,
у еще более стерильных - на зиготене.У самок млекопитающих первые стадии мейоза происходят еще до рождения. Мейоз доходит до стадии диплотены, а потом останавливается. Мейотические деления происходят уже у взрослых самок. В каждый репродуктивный цикл финальные деления совершает одна, редко две клетки у женщин или пять-семь клеток у самок пунаре.
Вся эта сложная последовательность событий детально запрограммирована. Строго по плану хромосомы сходятся, опознают друг друга, спариваются, обмениваются участками, расходятся. Строго по расписанию цепи ДНК разрезаются, спариваются, разрезы залечиваются. Согласно расписанию в клетке включаются и выключаются гены, ответственные за синтез белков, необходимых для обеспечения этих процессов.
Все идет, как на хорошо отлаженном конвейере по сборке автомобилей. Разница в том, что здесь машина сама себя собирает, более того, она еще и контролирует качество сборки. Прежде, чем перейти на следующую стадию мейоза, клетка сама себя проверяет, особенно придирчиво на переходе от пахитены к диплотене. Все ли хромосомы спарены? Не осталось ли неспаренных участков, незалеченных разрывов ДНК? Если все в порядке - идем дальше. Если хоть что-то не так, мейотическая клетка, как истинный самурай, делает себе харакири - переходит в апоптоз.
Забавно наблюдать, как клетка решает, казалась бы, неразрешимый конфликт: как быть с неспаренными участками половых хромосом у гетерогаметного пола? С одной стороны, спарить X с Y по всей их длине невозможно: уж слишком велики районы, их различающие. С другой, никак нельзя отступить от правила: все клетки с неспаренными хромосомами дальше пахитены не пускать, а немедленно отправлять в апоптоз. Для одной пары хромосом нельзя делать исключение из правила, поскольку контролеры исключений не понимают. Но можно их обмануть: спрятать пару не полностью спаренных половых хромосом и сделать ее (и только ее) недоступной для контроля. Именно так и поступают самцы млекопитающих. На стадии пахитены пара XY заботливо укутывается защитными белками и выпихивается на периферию ядра, подальше от систем контроля. Те ее не видят и дают зеленый свет для перехода на следующую стадию.
Если вернуться к разнице в мейозе между самцами и самками, то она не только в расписании основных мейотических событий, но и в строгости контроля. На первый взгляд, у самок контроль качества гамет должен быть более строгим, чем у самцов. Действительно, самки продуцируют относительно мало ооцитов. Именно самка вынашивает плод. Она должна быть уверена в качестве ооцита, из которого этот плод развивается. Если ооцит некачественный, долгое время беременности и ресурсы на развитие плода будут потрачены зря. А самцу - что? Его дурацкое дело нехитрое: наштамповать миллионы сперматозоидов, и если хоть десяток из них оплодотворит яйцеклетки, можно считать, что родительский долг исполнен.
На деле же все оказывается как раз наоборот. Контроль качества более строг при производстве мужских гамет. В зрелых ооцитах обнаруживается гораздо более высокий процент хромосомных аберраций, чем в сперматозоидах. У самцов-мутантов по мейотическим генам мейоз останавливается на стадии пахитены или еще раньше. У самок с теми же самыми мутациями мейоз доходит до конца и образуются зрелые ооциты. В ряде случае эти ооциты способны к оплодотворению и нормальному развитию.
Не в этом ли секрет правила Холдейна для млекопитающих? Если у гетерогаметного пола (самцов) мейотический контроль более суров, чем у гомогаметного (самок), то у гибридов-самцов он (контроль) будет браковать проблемные половые клетки и тем самым доводить этих самцов до полной стерильности.
Глава 5. Отчего одни гибриды стерильные, а другие еще стерильней
Теперь, когда мы разобрались с мейозом у нормальных фертильных самцов, самое время вернуться к стерильным гибридным самцам. Отчего и как у них возникают проблемные половые клетки, что в них не нравится контролерам, и когда эти клетки делают себе харакири?
У гибридов первого поколения между 30- и 28-хромосомными пунаре было 29 хромосом. Мейоз у них начинался нормально. Гомологичные хромосомы успешно находили друг друга, а хромосома X нашла хромосому Y. Каждая из 24 аутосом нашла себе пару. А оставшиеся три хромосомы образовали тройку: с одной стороны длинная хромосома, полученная от 28-хромосомного родителя, а с другой - две покороче, которые пришли от 30-хромосомного родителя. При этом каждая хромосома встала на свое место. Только их концы не смогли спариться с длинным партнером. Поэтому в одних клетках они спарились друг с другом, а в других остались неспаренными (рис.6). А за исключением этих мелких проблем, все было нормально. И тем не менее, ни одна половая клетка у самцов гибридов дальше этой стадии не прошла. Контролеры не пустили, и мейоз закончился на стадии пахитены. Неужели все из-за не спаренных или неверно спаренных концов хромосом?
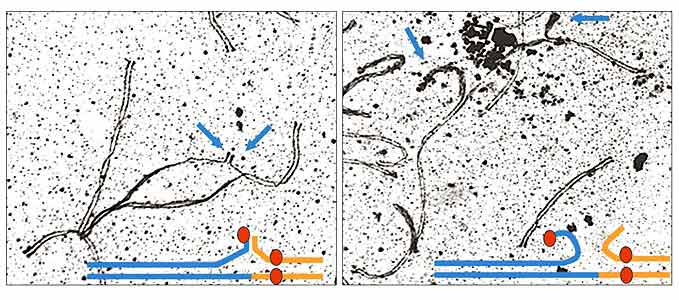
Рис. 6. В мейозе у гибридов перестроенная хромосома спаривается со своими короткими гомологами.
Вокруг точки слияния спаривание нарушено.
Концы коротких хромосом спариваются друг с другом (справа) или остаются не спаренными.Непохоже. Дело в том, что среди потомков от скрещивания гибридных самок с 30-хромосомными самцами были такие, у которых мейоз остановился на пахитене. А ведь они имели 30 хромосом, которые четко разобрались по парам и спарились полностью. И тем не менее, контроль не прошли.
Кроме того, у них были братья (те самые, что стерильней стерильных!). У них тоже было по 30 хромосом. Каждая должна была легко найти себе пару. Могла, да не нашла. Мейоз у этих братьев-пунаре доходил до зиготены. А далее, как сказал в аналогичной ситуации принц Гамлет, - молчание. Ни одна клетка пахитены не достигла. Все погибли.
Глава 6. Как наследуется гибридная стерильность у пунаре
Итак, было пять братьев пунаре. У всех по 30 хромосом (рис.4). Один был фертильный. Мейоз у него шел до конца и заканчивался формированием миллионов нормальных сперматозоидов. Двое других были стерильными. У них хромосомы спаривались, но мейоз остановился. А оставшиеся двое были еще стерильней. Хромосомы их не спаривались вовсе, а мейоз заканчивался едва начавшись (см. рис.5). Итого, мы имеем три фенотипа - норма, стерильные и еще стерильней.
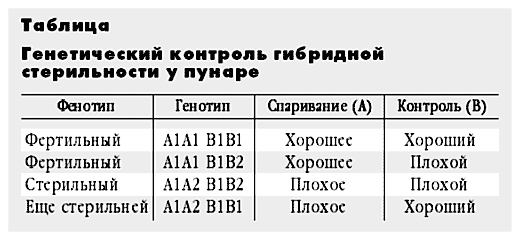
В генетике это называется расщеплением. Мы знаем, что если в потомстве от возвратного скрещивания есть три фенотипа, значит, в их контроль вовлечено как минимум два гена. Допустим, что один из них (А) контролирует спаривание хромосом, а другой (В) - контроль этого спаривания. Прадед, дед и отец наших братьев пунаре имели по 30 хромосом и были гомозиготами: А1А1, В1В1. Их прабабка имела 28 хромосом и была гомозиготой: А2А2, В2В2. У всех этих предков мейоз шел нормально. Их бабка была гетерозиготой по обоим генам А1А2, В1В2. У нее и спаривание было не слишком хорошим, и контроль качества не очень строгим. Но это ей не помешало размножиться, поскольку она была самкой. А у самок, как мы с вами уже выяснили, контроль всегда не строгий. Генотип матери братьев пунаре мы установить не можем, но допустим, что она, как ее мать, была гетерозиготой по обоим генам А1А2, В1В2.
Теперь разберем возможные генотипы ее потомков. При хорошем спаривании даже хороший контроль не страшен, а плохой уж и подавно. Самцы с генотипами А1А1, В1В1 и А1А1, В1В2 должны быть фертильны. У гетерозигот по обоим генам А1А2, В1В2 и спаривание идет плохо, и контроль плохой. Но поскольку они самцы, то все же не такой плохой, как у гетерозиготных самок. То, что у их матери и бабки доходило до конца мейоза, у них еле-еле добирается до середины. А дальше - конец. Они - стерильны. И, наконец, последний вариант А1А2, В1В1Ц спаривание плохое, а контроль такой хороший, что бракует клетки с неспаренными хромосомами в самом начале мейоза (табл.).
Эта схема выглядит вполне логично. Она удовлетворительно объясняет то, что мы наблюдаем у гибридов. Но как могла возникнуть в эволюции такая система гибридной стерильности?
Глава 7. Как возникала гибридная стерильность у пунаре
Отвечая на этот вопрос, мы должны исходить из того, что все три популяции пунаре, представителей которых мы скрещивали в лаборатории, имели общих предков. Эти предки, скорее всего, жили на юго-западе Бразилии, потому что именно там сохранились пунаре с наиболее примитивным 34-хромосомным кариотипом. Затем они расселились на север и восток. По мере расселения они теряли контакт друг с другом. Поток генов между отдельными популяциями ослабевал и наконец прекратился вовсе. После установления географической изоляции каждая популяция пошла своим путем эволюции. В разных популяциях возникали, распространялись и фиксировались разные мутации - генные и хромосомные. Все это происходило при одном условии - ни одна из этих мутаций в гетерозиготе не вызывала стерильности ее носителей. Иначе она не распространилась бы и не зафиксировалась.
Как при этом условии могли зафиксироваться мутации генов, контролирующих такие фундаментальные процессы, как спаривание хромосом и контроль качества половых клеток?
Рассмотрим сначала процесс спаривания. Вы помните, что он базируется на сравнении штрих-кодов гомологичных хромосом и на определении гомологий между отдельными участками ДНК. Механизмы распознавания не замечают гетерозотности по одной мутации. Спаривание наступает, если различия между гомологами не превышают определенный предел. Каждая популяция может постепенно меняться генетически, фиксируя одну мутацию за другой, без угрозы превысить этот предел. Но разные популяции накапливают разные мутации. За многие поколения независимой эволюции различия в последовательностях ДНК и в штрих-кодах их хромосом достигают того предела, когда эти хромосомы, встретившись у гибридов, с трудом узнают друг друга.
Точно так же постепенно и независимо друг от друга эволюционируют механизмы контроля качества мейотических клеток. В каждой популяции по-разному меняется расписание мейоза - продолжительность его отдельных стадий. Меняются метки на хромосомах, которые позволяют контролерам отличать качественное спаривание от некачественного. Когда мы сводим вместе в гибридных клетках эти далеко разошедшиеся системы контроля, они дают сбои.
В свете этих рассуждений нам следует переписать схему генетической детерминации гибридной стерильности у зверя пунаре. Мы описали ее в терминах двух пар генов - А и В (это, конечно, сильно упрощено). Скорее всего, нам следует говорить о двух генетических системах, каждая из которых включает в себя много генов. И популяции, из которых мы брали родителей для наших гибридов, различаются по многим генам, входящим в эти системы. Еще популяции или уже виды?
Глава 8. Сколько видов среди зверей пунаре
Вот мы и вернулись к вопросу, заданному в самом начале этой истории. Так сколько же видов среди пунаре? Один или несколько? В школьных учебниках описано несколько критериев вида. Попробуем их применить к пунаре. По морфологическому критерию его разные популяции явно не тянут на виды. Разницы между ними никакой. По экологическому - может, и тянут. Уж очень сильно отличаются условия, в которых живут наши популяции: в болотах Пантанала, в саваннах центральной Бразилии, на горных склонах Сьерры Капибары. По цитогенетическому критерию они вполне подходят под ранг видов - все три популяции различаются по кариотипам.
Самой важный из критериев - репродуктивная изоляция. Мы показали, что самцы-гибриды между разными популяциями стерильны. Но самки-то фертильны. Так есть ли между этими популяциями репродуктивная изоляция или нет? Есть, но неполная. Так виды это или не виды? Почти виды, но еще не совсем. К сожалению, мы пока ничего не знаем, что происходит на географических границах между популяциями. Соприкасаются ли ареалы разных популяций или нет? Если да, то образуются ли гибриды в природе? Неизвестно. Но очевидно, что звери пунаре уже далеко зашли по пути к видообразованию. А что будет с ними дальше - покажет время.Благодарности.
Цитогенетический и гистологический анализ проводился в Новосибирске. За помощь в приготовлении гистологических препаратов мы благодарны А.И.Железовой и Л.А.Чугаевой.
Работа была выполнена при поддержке грантов Бразильского совета по науке и технологиям (CNPq), Фонда Освальдо Круза (FIOCRUZ), и РФФИ (01-04-48875 и 04-04-48024).
Литература
1. Поляков А.В., Бородин П.М. Хромосомный "портрет" бурозубки на фоне ледников // Природа. 2001. No.1. С.34-40.
2. Бородин П.М., Рогачева М.Б., Ода С.И. Домовая землеройка на пути к видообразованию // Природа. 2002. No.9. С.1-11.
3. Богданов В.Ф. Изменчивость и эволюция мейоза // Генетика. 2003. Т.39. С.453-473.